1 Introduction
The Haynie site (5MT1905) is a multicomponent ancestral Pueblo archaeological site, located in southwestern Colorado, occupied between roughly A.D. 800 and 1200 (Throgmorton et al. 2022). It includes two Chaco period Great Houses. Material recovered from Haynie represents an unparalleled way to examine how Chaco outliers formed, were sustained, and integrated within the larger Chaco world.
The Lakeview Community encompasses the Haynie site and two other Great Houses, all of which are located within 1 km of each other. The additional Great Houses are Ida Jean (5MT4126) and Wallace Ruin (5MT6970). Studying the Lakeview Community is the purview of the Northern Chaco Outliers Project. Crow Canyon initiated this project in 2016 (Ryan 2016), and it has four research domains: human-environment relationships, social inequality, the role of public architecture in Chaco Outlier communities, and identity formation. The excavation and analysis of material culture from Haynie is one of the prime ways these research domains will be addressed.
Studying archaeofaunal remains is an integral part of the Northern Chaco Outliers Project, as this class of material fits within each research domain. How humans interact with animals represents a significant portion of how they generally interact with environments. Animal foods can also help detect social equality or inequality (Badenhorst et al. 2019; Muir and Driver 2002), are integral to the use of public architecture (Potter 2000; Potter and Ortman 2004), and can be an important identity marker (Potter 2004; Twiss 2007). Further, animal parts were used for purposes other than food that can be applied to these research domains. Animal parts were used in the manufacture of tools and ornaments that were incorporated into rituals, ceremonies, and domestic activities (Brown 1967; Muir and Driver 2004; Olsen 1979, 1980; Payne 1991; Watson and Gleason 2016). The Haynie archaeofauna represents a way to directly understand how and why Chaco Outlier community members obtained and used animals.
This report, however, focuses on three areas that undergird these research domains: the identification, taphonomy, and representative sampling of the Haynie site archaeofauna. We detail how remains were identified, their condition, and how the diversity of animals recovered compares to various sites in the northern U.S. Southwest. In the identification section, we especially emphasize the research potential of artiodactyl and canid remains (Section 3). The taphonomy section emphasizes fragmentation, as its effects were conspicuous (Section 4). Finally, the section on sampling compares the structure of taxonomic diversity at Haynie and other sites (Section 5). This section is especially essential considering that representative sampling underlies almost every facet of zooarchaeological interpretation (Grayson 1984, 1978, 1981; Lyman 2008). Overall, the goal of this report is to document the quality of zooarchaeological data produced from August 2021 to August 2022. Tracking data quality is fundamental to understanding the range of research applications for the Haynie site archaeofauna. Such a focus highlights strengths and weaknesses of these data, which will help legitimately address the research domains of the Northern Chaco Outliers Project in the future.
2 Materials and Methods
We are the sole analysts of the data described herein. Currently, Jonathan Dombrosky has approximately 11 years of experience with archaeofaunal analysis, and he analyzed specimens from August 2021 to August 2022. Eric Gilmore has approximately 3 years of experience with archaeofaunal analysis, and he analyzed specimens as a Crow Canyon Zooarchaeology Intern from May 2022 to July 2022. Jonathan Dombrosky and Eric Gilmore are respectively referred to as Analyst 1 and Analyst 2 in all subsequent interanalyst comparisons.
The main comparative collection used for the analysis of the Haynie site archaeofauna is housed in Crow Canyon Archaeological Center’s Laboratory. Three cottontail (Sylvilagus spp.) specimens and one black-tailed jackrabbit (Lepus californicus) specimen were loaned from the Laboratory of Zooarchaeology at the University of North Texas. We took specimens that were difficult-to-identify to the Museum of Southwestern Biology’s Division of Mammals and Division of Birds located in Albuquerque, New Mexico. Various osteological guides, manuals, atlases, and keys aided identification. Several publications assisted with the identification of mammal remains (Adams and Crabtree 2012; Chavez 2008; Gilbert 1980; Hillson 1986, 1996; Jacobson 2003; Olsen 1964; Smart 2009). Bird remains were identified with other works (Cohen and Serjeantson 1996; Gilbert et al. 1981; Hargrave and Emslie 1979; Olsen 1979). Some consulted references helped identify both avian and mammalian remains (Broughton and Miller 2016; Elbroch 2006). Yet other works verified nonhuman from human remains (Baker et al. 2005; France 2009; White et al. 2012).
We followed identification protocols explicitly designed to enhance data quality (Driver 1992, 2011; Wolverton 2013; Wolverton and Nagaoka 2018), and used the coding system by Driver (2006). Briefly, analysts adopted a conservative approach to identifying zooarchaeological specimens at the Haynie site. It is an almost impossible task for analysts to understand how all diagnostic skeletal criteria change through time, among species, within different age classes, between sex, and across geographic areas on a fragment-by-fragment basis. It has been argued that identifications become less taxonomically specific when analysts have more experience, greater access to diverse comparative materials, and a specific focus on data quality (Gobalet 2001; Lyman 2002; Wolverton and Nagaoka 2018). This lack of taxonomic specificity likely increases identification accuracy in situations where assemblages contain an abundance of fragmented remains from closely related taxa.
We use the Number of Identified Specimens (NISP) to report taxonomic abundance, and this quantitative unit is a tally of all archaeofaunal specimens within a given taxonomic classification. NISP is the most basic quantitative unit from which most others are derived, such as the Minimum Number of Individuals (MNI). NISP is preferred because it is often highly correlated with measures like MNI. It is also devoid of errors in additive calculation that plague minimum number units (Grayson 1984; Lyman 2008). We also rely on a non-standard unit called the Number of Unique Identifications (NUIDs) to estimate taxonomic diversity (Section 5).
All statistical analyses and figures were produced with R version 4.2.0 (R Core Team 2022). Our statistical analyses are structured with tidyverse packages and syntax (Wickham et al. 2019). All graphs were produced with ggplot2 (Wickham 2010). We built predictive taxonomic and taphonomic models using a supervised learning workflow (Hastie et al. 2009; James et al. 2013; Kuhn and Johnson 2013); this included using the tidymodels metapackage to split our data and implement basic model features (Kuhn and Silge 2022). We rely on Linear Discriminant Analysis (LDA) and logistic regression as the engines for our predictive models. LDA is a procedure designed to maximize the linear separation of multiple classes based on given predictors (Kachigan 1991), such as length-based measurements. Logistic regression is a popular modeling engine designed for binary classification (Kuhn and Johnson 2013, 282).
3 Identified Taxa
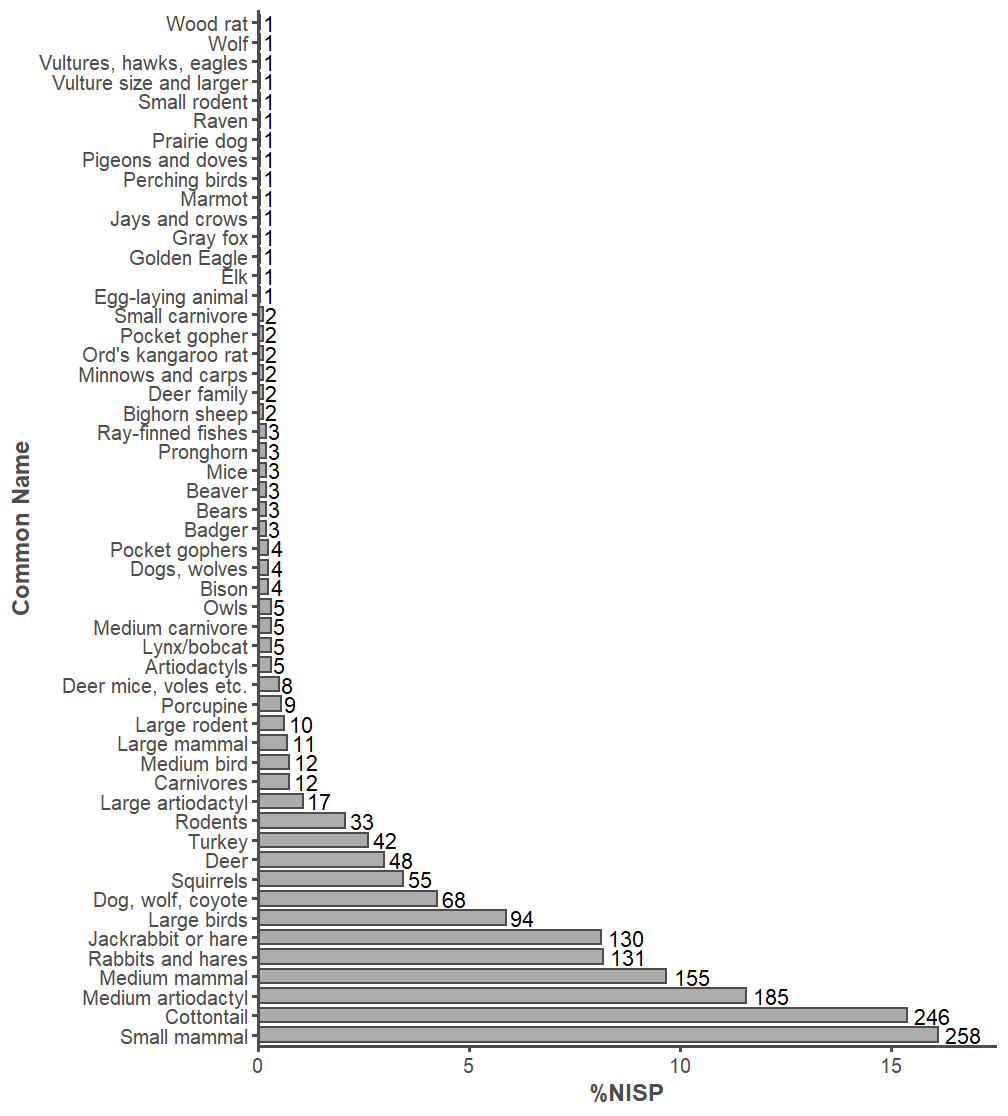
There are 1601 identifiable and 4281 unidentifiable specimens so far in the Haynie archaeofaunal assemblage. This leads to an identification rate of 27.22%, which is low and likely driven by fragmentation issues (Section 4). Analyst 1 analyzed 72.58% of the assemblage, and Analyst 2 analyzed 27.42%. There are 3 classes of animals present: Mammalia (mammals), Aves (birds), and Actinopterygii (ray-finned fishes). There are 4 orders of mammals present, 6 orders of birds, and 1 order of ray-finned fishes. In total, we used 54 identification types (Figure 1).
3.1 Mammalia (n = 1435)
3.1.1 Lagomorpha (n = 507)
Cottontails (Sylvilagus spp.) and jackrabbits (Lepus spp.) are the main taxa in the order Lagomorpha at the Haynie site. Lagomorphs are currently the most abundant animals identified, comprising 31.67% of the identified specimens. It has been hypothesized that populations of larger-bodied jackrabbits decreased through time in the central Mesa Verde region, and that this decrease is likely due to human overhunting (Driver 2002; Ellyson 2014). Thus, the ratio of cottontails to jackrabbits—commonly referred to as the Lagomorph Index—is a basic quantitative unit of general interest in the area, and in the broader U.S. Southwest (Driver and Woiderski 2008). The Lagomorph Index at the Haynie site is 0.65, which indicates a fairly equal relationship between the abundance of cottontails and jackrabbits. This number is similar to the Lagomorph Index from some Pueblo II components of Shields Pueblo (5MT3807). Shields serves as an important point of comparison—here and in subsequent analyses—considering that it and Haynie both have similar site features (i.e., Great Houses) and general occupation histories (Rawlings 2006).
Garden hunting is an important subsistence practice to consider at the Haynie site, and the moderate Lagomorph Index value is also interesting in this regard. Researchers argue that higher ratios of cottontails to jackrabbits indicates a higher reliance on garden hunting in the central Mesa Verde region (Driver 2011). What could the more even relationship of cottontail and jackrabbit abundance indicate about garden hunting at the Haynie site? Future work using stable isotope analysis might help shed light on whether garden hunting was prevalent at the site, and it also might help describe the relationship between the Lagomorph Index and garden hunting in general.
3.1.2 Artiodactyla (n = 267)
Even-toed hoofed animals make up the order Artiodactyla, and they are 16.68% of identified specimens at Haynie. There is substantial zooarchaeological evidence that indicates artiodactyls—mostly mule deer (Odocoileus hemionus), pronghorn (Antilocapra americana), and bighorn sheep (Ovis canadensis)—were severely overhunted in the central Mesa Verde region (Badenhorst and Driver 2009). Some models suggest that deer populations might have been so low that any observable deer was immediately hunted after A.D. 1000 (Bocinsky et al. 2012). For this reason, the Artiodactyl Index is another basic quantitative unit of general interest in the region and in the larger northern U.S. Southwest. It measures the ratio of large-bodied artiodactyls to small-bodied lagomorphs in an archaeofaunal assemblage (Broughton et al. 2011). The current Artiodactyl Index at Haynie is 0.34. This value may seem somewhat low but the majority of Pueblo I and Pueblo II sites in the central Mesa Verde region have Artiodactyl Index values between 0.00 and 0.20 (Badenhorst and Driver 2009, Table 5). The value at Haynie is moderate.
Large artiodactyls—elk (Cervus canadensis) and bison (Bison bison)—are notable parts of the Haynie archaeofaunal assemblage. These specimens comprise 8.24% of artiodactyls. There are 4 bison specimens firmly identified: a complete first phalanx (Figure 2), a proximal metatarsal (Figure 3), a distal humerus, and a spinous process fragment from a thoracic vertebra. There are also other specimens that compared favorably to bison but did not retain enough morphological characteristics for firm identification: a thoracic vertebra neural arch fragment and a rib shaft fragment (Figure 4).
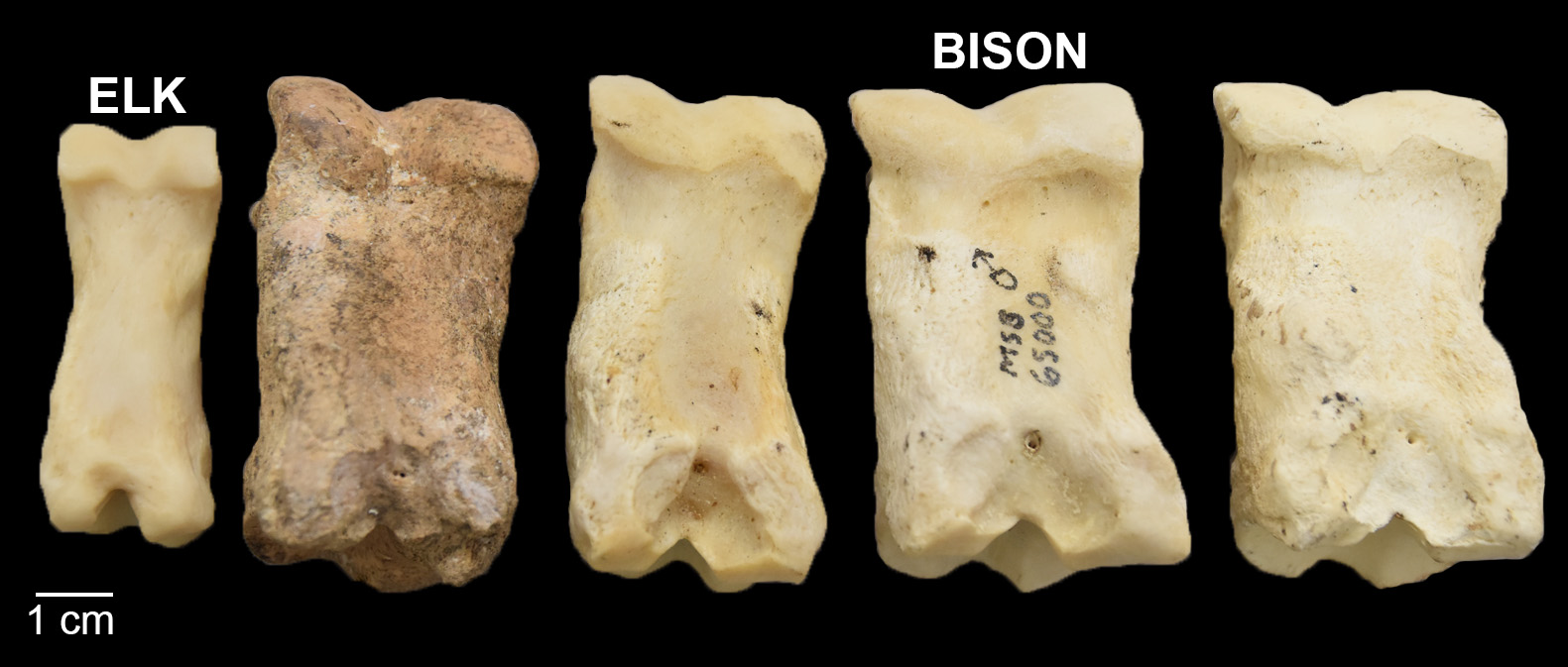

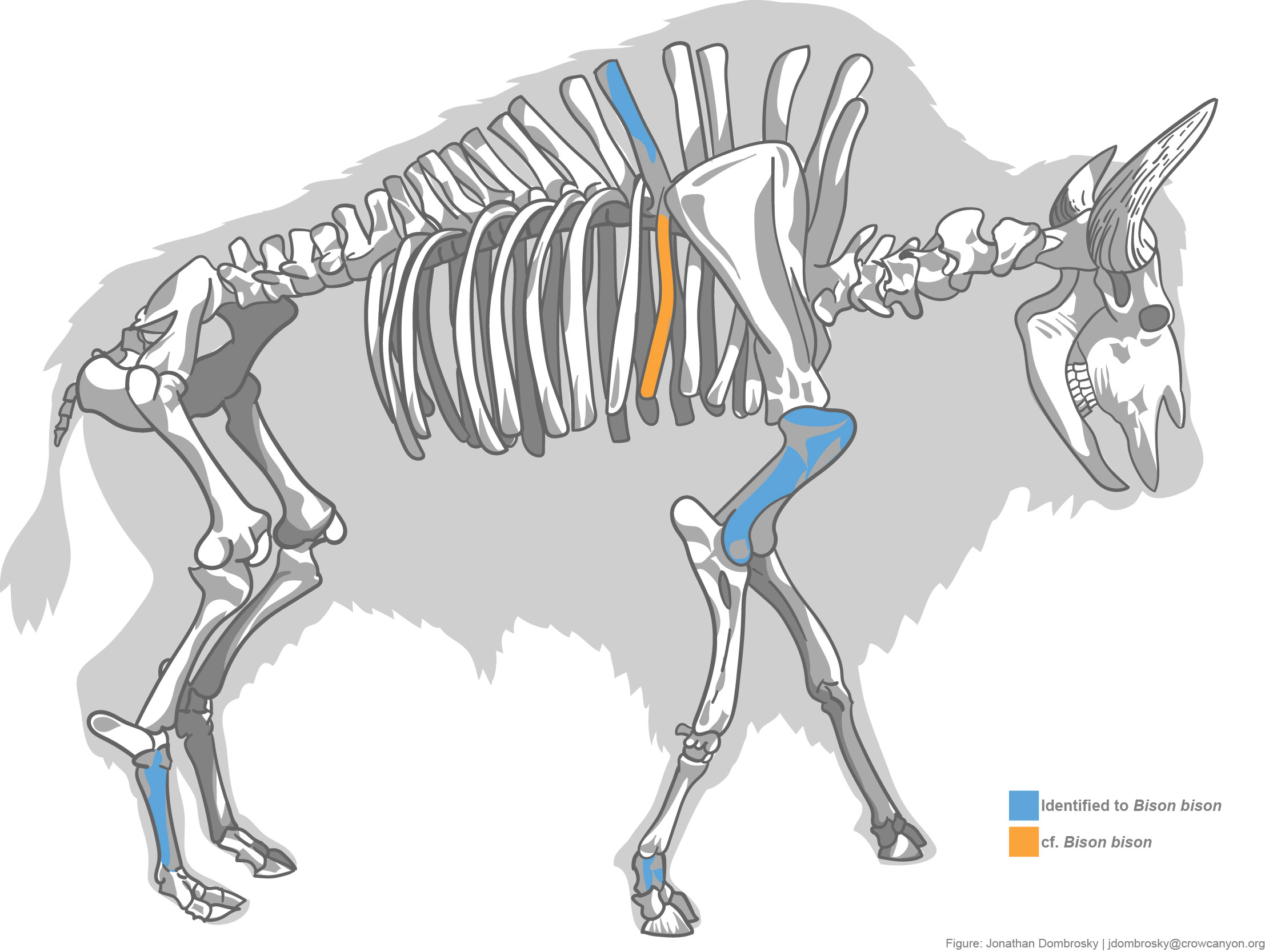
Bison remains are rare in the central Mesa Verde region. Only 4 specimens have been identified by Crow Canyon analysts. Two specimens, an ulna and rib fragment, were identified from Albert Porter (5MT123) by Badenhorst and Driver (2015). The other two, a complete first phalanx and a fragmented second phalanx, were recently identified by Cates (2020) from Greenstone Pueblo (5MT6970), which is located close to the Haynie site. Bison are also reported from the Badger House community in Mesa Verde National Park (Hayes and Lancaster 1975; Martin et al. 2017). The presence of bison remains at Haynie leads to a number of questions about how resources were obtained. Was the bison hunted locally, non-locally, or was it traded in? Skeletal element representation indicates elements of various utility are present, which might suggest bison parts were taken in bulk close to the Haynie site (sensu Binford 1978). Analyzing medium artiodactyls may be another indicator of the prevalence of local versus non-local hunting practices.
Specifically, it might help to assess the number of mule deer relative to pronghorn at the site. More mule deer may suggest that hunting locally was common, while more pronghorn would suggest non-local hunting practices prevailed. Given the habitat preferences of these two taxa, mule deer are assumed to have been procured near the Lakeview Community while pronghorn would require further travel to the south. Such a comparison, however, relies on accurate identification of mule deer and pronghorn specimens.
Analyst 2 measured the tarsals of 17 mule deer and 30 pronghorn from the Museum of Southwestern Biology’s Division of Mammals to build an accurate predictive taxonomic model. We used Linear Discriminant Analysis (LDA) as the engine, where it classifies a set of archaeological tarsal measurements as pronghorn or mule deer. Separate LDAs were used to create more accurate models for the astragalus, calcaneus, and cubonavicular. The astragalus model is 97% accurate, the calcaneus model 95%, and the cubonavicular is 93% accurate. The models identified 1 pronghorn and 12 mule deer when applied to Haynie, which indicates substantially more mule deer than pronghorn tarsals in the assemblage (Figure 5).
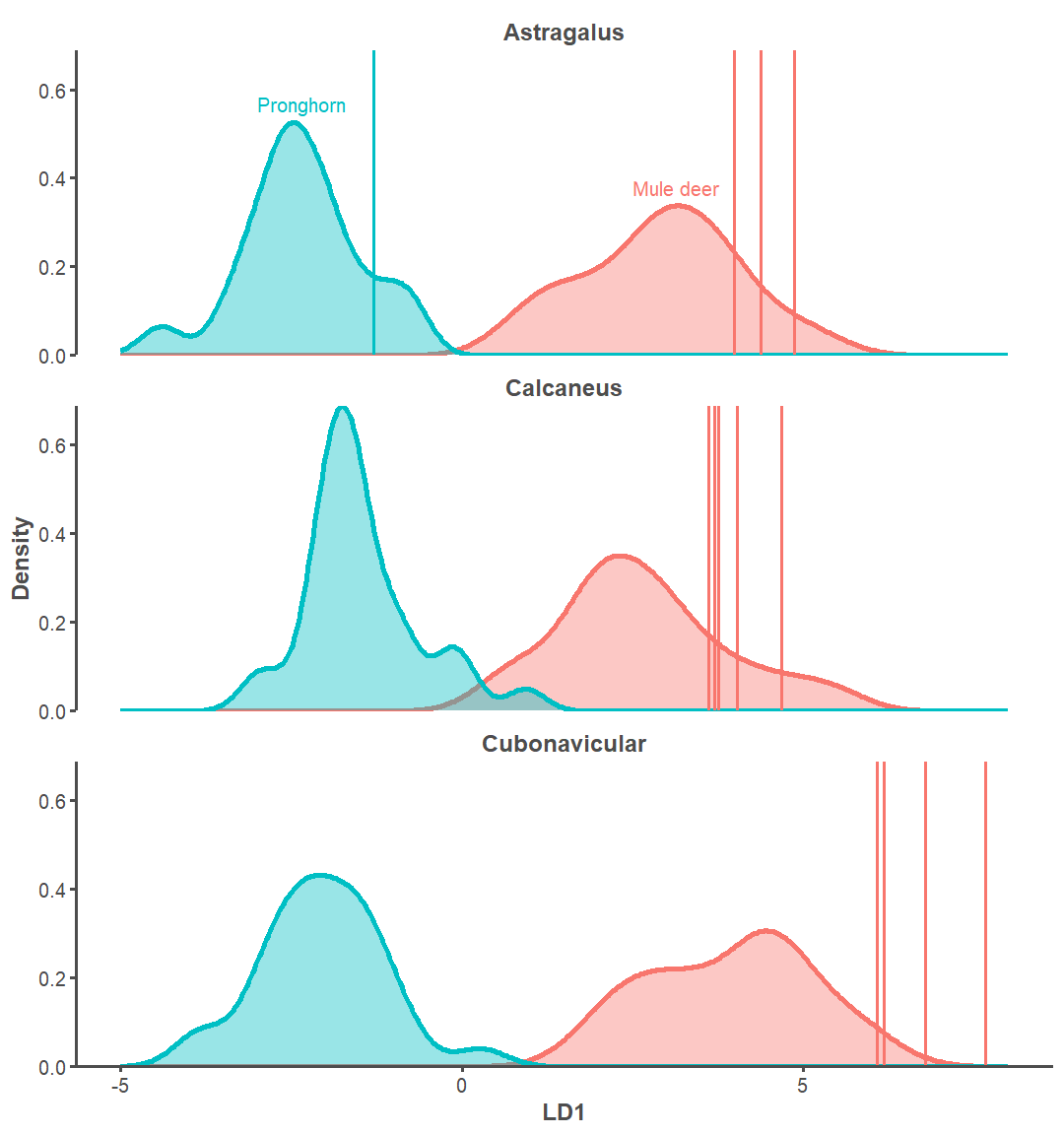
Interestingly, Analysts 1 and 2 did not originally identify pronghorn from this sample of tarsals (Figure 6). The predictive model only assigns elements to mule deer or pronghorn and the medium artiodactyl classification is only used with visual identifications. The model developed here improved taxonomic resolution at Haynie.
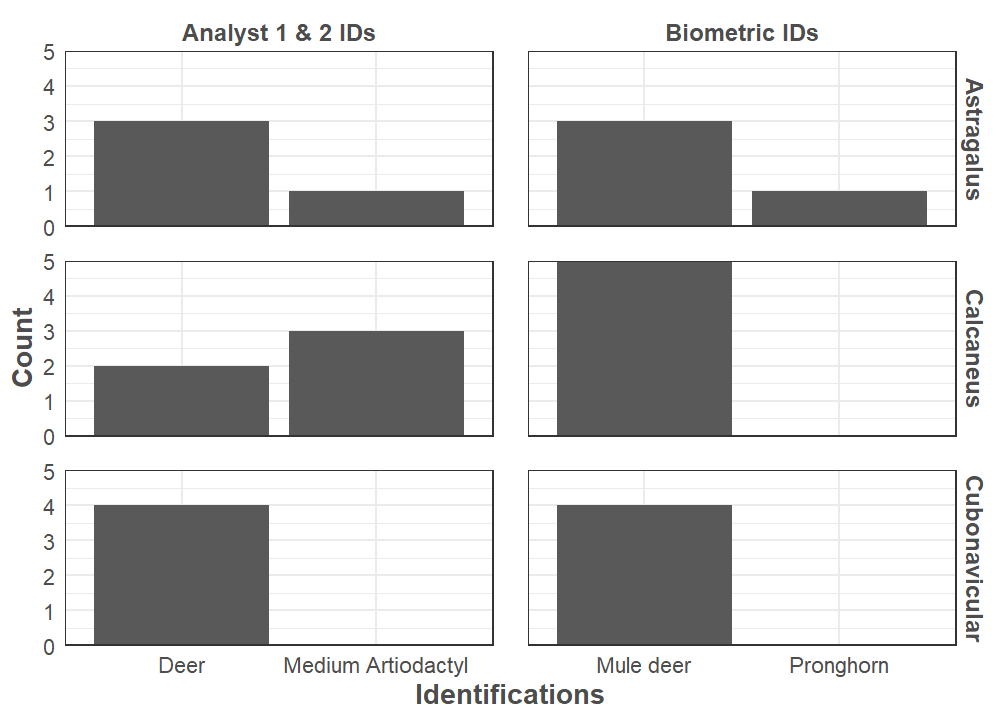
These results are one line of preliminary evidence suggesting that animal procurement was likely mostly local at the Haynie site. The original visual identifications indicated animals were solely procured locally, as the medium artiodactyl identification group kept the possibility of non-local procurement vague. Biometric analysis suggests that local procurement is still likely the predominate method, but that some long distance procurement did indeed occur. Applying these conclusions to the presence of bison remains suggests that bison were procured close to the Lakeview Community, but that long distance hunting and trade cannot be ruled out yet. Further analyses—including radiocarbon, stable isotope, and ancient DNA—may help address this issue.
3.1.3 Small Mammal (n = 258)
This identification group includes those mammals jackrabbit size and smaller. This non-standard identification includes all small mammal specimens lacking morphological features required for more specific taxonomic levels. Consequentially, it has a high likelihood of incorporating many lagomorph specimens since they are exceedingly abundant in southwestern archaeofaunas. It is crucial to incorporate this identification group in any rigorous comparison of size-based abundance indices through time, between areas of the Haynie site, or between sites. It should be incorporated in a sensitivity analysis to assess whether or not it impacts final interpretation of measures like the Lagomorph Index.
3.1.4 Medium Mammal (n = 155)
Mammals larger than a jackrabbit and up to deer size are considered medium mammals. There is a high probability this identification group incorporates many artiodactyl specimens since they are one of the most common medium mammals in southwestern archaeofaunas. It should be incorporated in sensitivity analyses that rigorously assess final conclusions relying on size-based abundance index values, such as the Artiodactyl Index.
3.1.5 Rodentia (n = 133)
Rodents are 8.31% of identified specimens, which is a small component of the overall assemblage. The ratio of sciurids to rodents can help gauge the impact of intrusive species. Members of the squirrel family (Sciuridae) are notorious intruders of archaeological deposits, especially prairie dogs (Cynomys spp.). The majority of prairie dog skeletal fragments are identified to the family level, as it is extremely difficult to skeletally distinguish prairie dogs from ground squirrels. This means that the Sciuridae identification has the highest potential for accumulating prairie dog specimens. And, indeed, we can confirm that the vast majority of sciurid specimens compare favorably to prairie dogs. Sciurids make up 42.86% (n = 57) of the rodent assemblage. Given that rodents comprise a small portion of the overall assemblage, it does not appear that rodent intrusions pose a significant problem for interpretation at the site. We will, however, closely track the taxonomic composition of rodents as the Northern Chaco Outliers project progresses. Importantly, rodents could have been actively hunted (Badenhorst et al. In press). One way to disentangle intrusive from non-intrusive rodents is through radiocarbon dating (Guiry et al. 2021). This could prove useful if prairie dog specimens preclude clear interpretation of the Haynie archaeofaunal assemblage in the future.
Large rodents, those larger than a woodrat (Neotoma spp.), comprise 17.29% of the total rodents currently identified. Beaver (Castor canadensis) and porcupine (Erethizon dorsatum) specimens are the most notable. Beavers prefer aquatic habitats while porcupines take refuge in trees along active floodplains (Baker and Hill 2003; Roze and Ilse 2003). The presence of these species might suggest hunting activities that were focused in riparian habitat close to the site.
3.1.6 Carnivora (n = 104)
Carnivores are a small portion of currently identified specimens at the Haynie site (6.5%), but there are a number of specimens worth detailed attention.
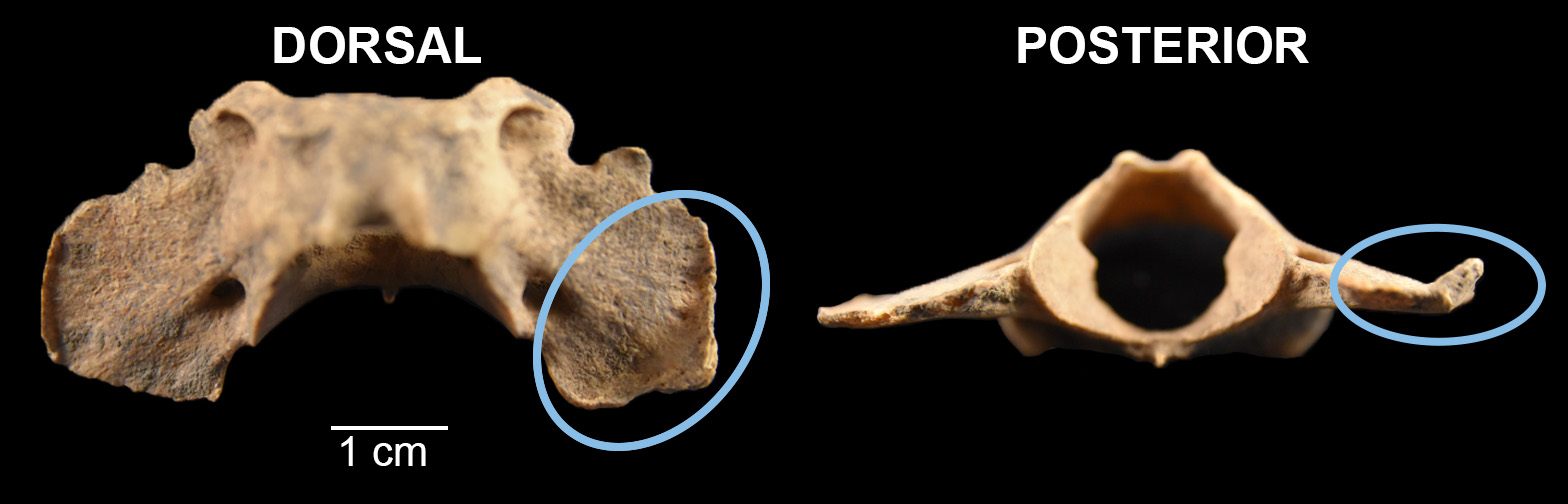
The first specimen is a wolf mandible (Canis lupus; Figure 7). It was recovered from the fill between two floors of a surface room in Structure 1026/1042 (see Throgmorton et al. 2022, fig. 6). It likely dates somewhere between A.D. 940 and 1040 based on associated artifacts. The second set of specimens are from the articulated remains of what is likely a small domestic dog (Canis familiaris). The upper half of this individual was uncovered from the floor of a pitstructure, Structure 1047 (Throgmorton et al. 2022, fig. 6). Its lower half was deposited outside of the unit. This individual likely dates to around the A.D. 1050s based on associated artifacts. Canids are known components of dedicatory offerings for room closing rituals (Hill 2000), and are recovered from room surfaces throughout the Pecos chronology (Burger 2021; Frisbie 1967; Lang and Harris 1984; Strand 1998). Interestingly, this individual was also found in association with an articulated turkey, which is another noted pattern (Burger 2021; Hill 2000). All analyses were conducted in situ for this individual dog given its cultural relevance. Exposed specimens (Figure 8) were identified and various mandibular measures taken.
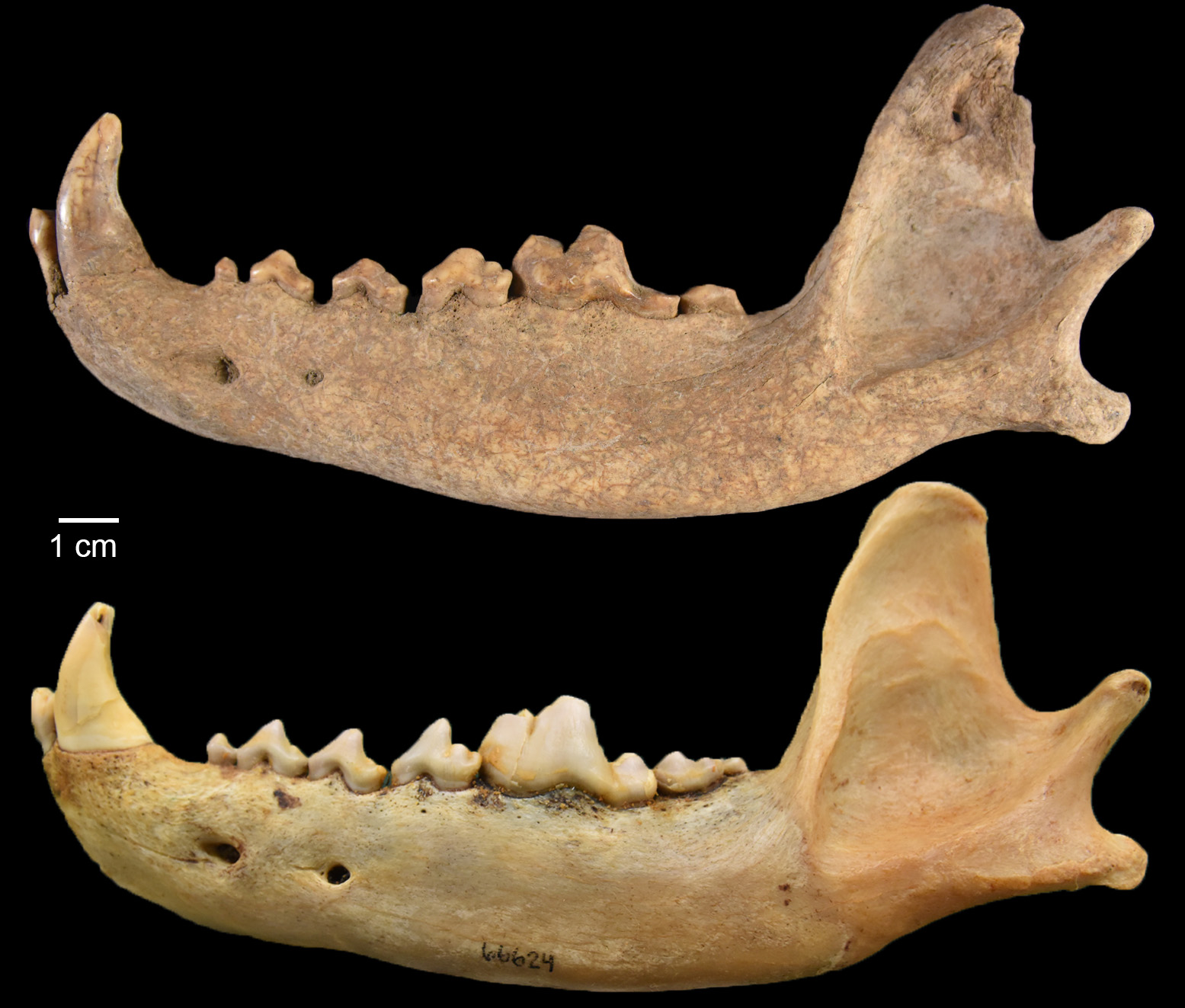
These specimens are likely Canis lupus and Canis familiaris based on visual inspection and comparison. We, however, wanted to know the level of confidence in our identifications considering the large amount of morphological overlap in members of the family Canidae (Wayne et al. 1997). Developing confidence-based taxonomic identification is a critical future avenue for zooarchaeology (Lyman 2019). Luckily, Welker et al. (2021) provided a robust (n = 543) database of North American canid mandibular measures.
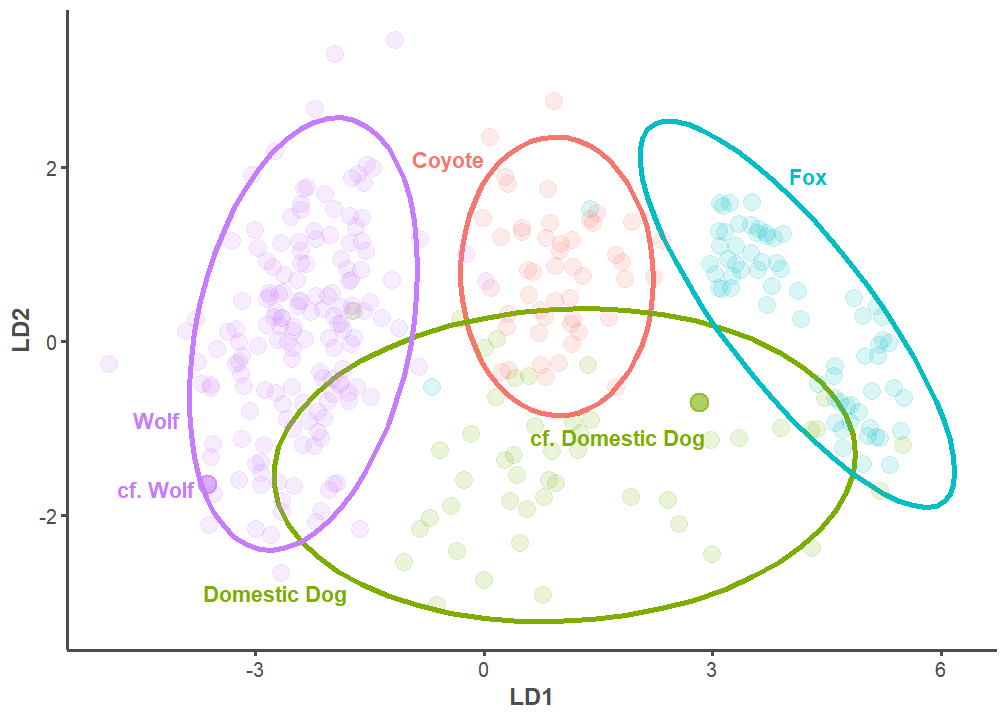
We built a model to classify canid mandible specimens from the Haynie site using this database. First, we isolated all species that can be found in the central Mesa Verde region: domestic dog, wolf, western coyote (Canis latrans), gray fox (Urocyon cinereoargenteus), kit fox (Vulpes macrotis), and red fox (Vulpes vulpes) specimens (n = 400). We then designed a model to classify domestic dogs, wolves, coyotes, and foxes (Urocyon sp. and Vulpes spp. were grouped together) by a set of five predictors: the length of p2, p3, p4, m1, and the height of the mandible behind m1 (see von den Driesch, Angela 1976, 60; Welker et al. 2021, 199). We trained the model on a random 80% split of the data. The model correctly classified 94.69% of the training dataset. We used 10-fold cross validation to evaluate model performance and gauge overfitting (Figure 9), and model performance was acceptable. We then applied it to the testing dataset, which consists of the remaining 20% of data from our initial split. We found that it correctly classified 88.75% of test data. This level of accuracy is considered generally high (Kuhn and Johnson 2013, 254).
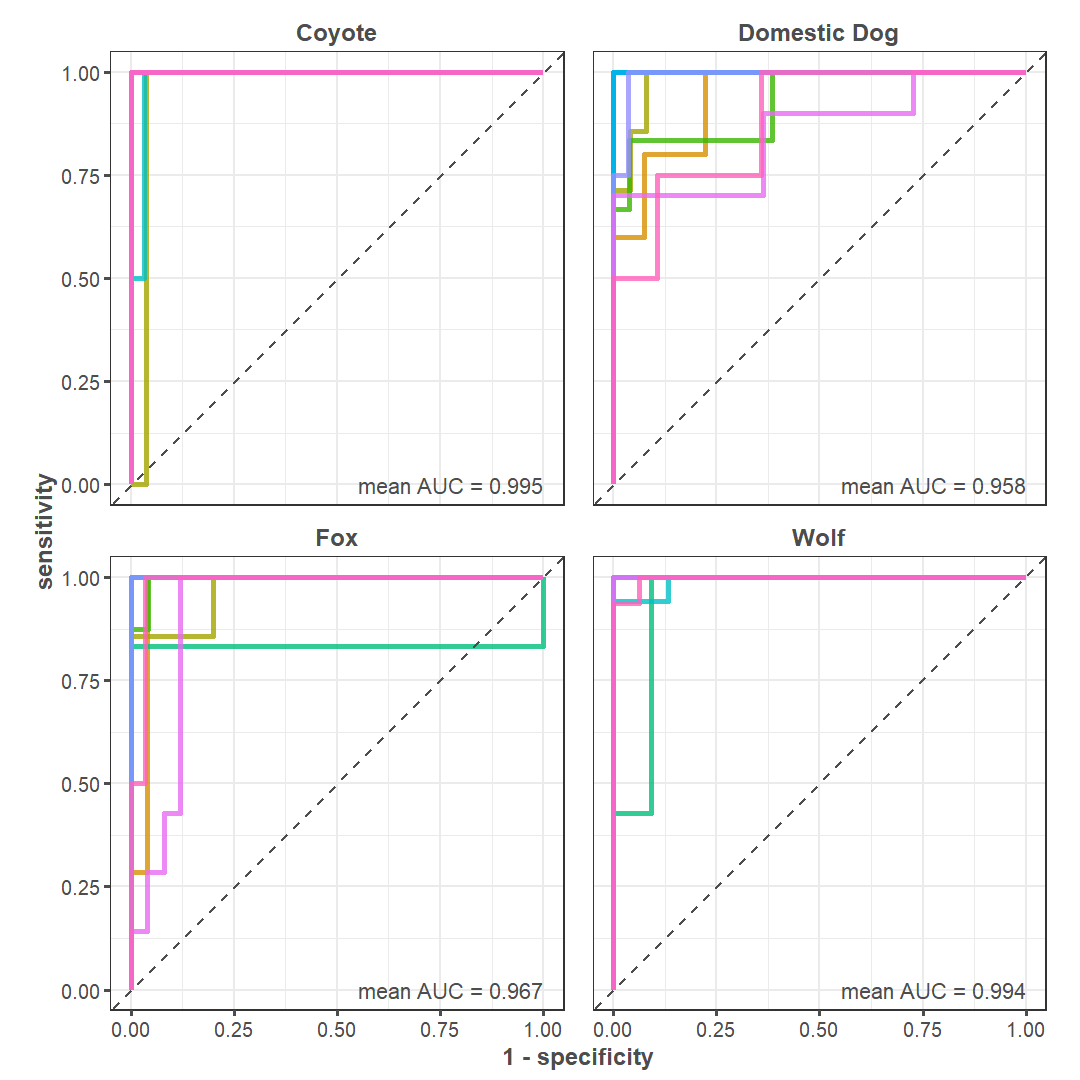
As anticipated by visual identification, the model predicts that the mandible specimens recovered from Structure 1026/1042 and 1047 respectively belong to wolf and domestic dog (Figure 10). But, returning to the question at hand, how confident can we be in these identifications? The posterior probability—or the likelihood of an identification occurring given the model—for the wolf specimen is 100%. In other words, the model is certain the specimen from Structure 1026/1042 is from a wolf. The posterior probability for the likely domestic dog specimen is slightly more complicated: there is a 44% likelihood the specimen is a domestic dog, 38% chance it is fox, and 18% probability it is coyote. It is unsurprising that the model has a more difficult time predicting domestic dogs (see Figure 9) given their extreme morphological plasticity. This concept is well-illustrated in Figure 10. Though there is a level of uncertainty, we feel confident in this model’s final classification of domestic dog. The cranium articulating with this mandible was not as gracile as a fox nor was it as elongated as a coyote. The depositional context also helps bolster the domestic dog identification, given that articulated domestic dogs are commonly recovered on room floors (Hill 2000).
The wolf specimen is also of particular interest because they are rarely recovered in the central Mesa Verde region. Crow Canyon zooarchaeologists have only identified four wolf specimens: a fragmented radius and complete atlas from Albert Porter along with a fragmented sphenoid and a femur with a healed fracture from Shields Pueblo (Badenhorst and Driver 2015; Rawlings and Driver 2015). This is not to say these are the only specimens in other assemblages, but wolf specimens identifiable beyond Canis sp. are extremely rare. This single specimen from Haynie represents an unprecedented way to study human-wolf relationships in the region and in the northern U.S. Southwest considering how complete it is. For instance, toothwear suggests this individual was old, likely somewhere between 8 and 10 years old following the guidelines provided by Gipson et al. (2000). It is also possible this individual could be greater than 12 years old considering the intense wear on the posterior portion of m1 (see Figure 7). This locus of intense wear is certainly interesting, but any definite interpretation must consider bite variation among individual gray wolves. Regardless, the mean age at death for wolves in the wild is approximately 3.76 years old, which suggests the Haynie specimen reached a particularly old age despite the range in toothwear estimates (Figure 11). Preliminary modeling also suggests the individual was a male. This particular model was 80.95% accurate with training data, but only 63.63% accurate with test data. These conclusions are certainly not definitive, but the male posterior probability for this specimen is 88.78%. Future work with this specimen will focus on strengthening models to predict sex, and may likely include analysis of radiocarbon, stable isotopes, and ancient DNA.
There are also interesting carnivore specimens beyond the wolf and individual domestic dog. One of the clearest signs of pathology comes from a bent transverse process on a Canis sp. atlas (Figure 12). Vertebral deformation is common in domestic dogs from ancestral Pueblo contexts (Monagle and Jones 2020), and this type of pathology could be indicative of mechanical compression. Potential causes for this type of compression include tethering or possibly even carrying a pack with a heavy load. However, natural variation in vertebral morphology as domestic dogs age should be assessed too.
Other carnivore specimens worth noting include a drilled bobcat (Lynx rufus) terminal phalanx, and a bear terminal phalanx (Figure 13). Human-carnivore relationships are an exciting area of future study at Haynie and will focus on how these animals were procured and cared for.

3.1.7 Large Mammal (n = 11)
The large mammal identification includes mammals larger than deer, and it includes specimens lacking morphological features required for more specific taxonomic levels. It is likely that it could incorporate large artiodactyls like elk and bison specimens. It should be incorporated in sensitivity analyses that rigorously assess conclusions based on these animals.
3.2 Aves (n = 160)
3.2.1 Large Birds (n = 94)
Birds larger than a mallard are considered large birds. This identification group is most likely dominated by Turkeys (Meleagris gallopavo) as they are one of the most frequent birds recovered from ancestral Pueblo sites. A high proportion of Turkey specimens are assigned to this group considering that there is considerable skeletal morphological overlap between Sandhill Crane (Grus canadensis) and Turkey (Hargrave and Emslie 1979). The Crow Canyon comparative collection does not, as of yet, include Sandhill Crane skeletal material.
3.2.2 Galliformes (n = 42)
All Galliformes specimens identified so far in the Haynie assemblage are Turkey, but it is surprising how few Turkey specimens there seem to be. For instance, Spielmann and Angstadt-Leto (1996) proposed the Turkey Index, which is the ratio of Turkeys to lagomorphs in an assemblage. The current Turkey Index for Haynie is 0.08, which is notably low. However, Driver (2002) proposed the Modified Turkey Index where the large bird identification group is included in the calculation. The current Modified Turkey Index value for Haynie is 0.23, which is within the normal range for Pueblo I/Pueblo II sites in the central Mesa Verde region (Badenhorst and Driver 2009). Also worth noting, there appears to be more turkey specimens yet to be identified. How Turkey husbandry was managed at the community-level is an essential future area of research for the Northern Chaco Outliers Project. This line of inquiry is one way to delve deeper into aspects of cooperation and identity at the Lakeview Community.
3.2.3 Medium Birds (n = 12)
Medium birds are considered larger than a robin and the size of a mallard or smaller. It is difficult to attribute the majority of this identification group to a single taxon, as it contains a variety of difficult-to-identify fragmented skeletal parts that could belong to numerous taxa.
3.2.4 Strigiformes (n = 5)
The owl specimens identified at the Haynie site are a terminal phalanx, second phalanx, first phalanx, femur, and ulna. Owl feathers are known to have been incorporated into dance paraphernalia and prayer sticks (Ladd 1963). It is, however, important to keep in mind that owls can be active taphonomic agents. Luckily, their signatures are well-known and include the presence of pellets, small mammal remains with little to no fragmentation, and visible signs of digestion on specimens (Andrews and Cook 1990; Fernández-Jalvo and Andrews 2016). Owls do not appear to be a taphonomic agent of concern at Haynie (Section 4).
3.2.5 Accipitriformes (n = 3)
Two specimens were identified as general members of the order Accipitriformes: one foot phalanx and one terminal phalanx that compared favorably to a Turkey Vulture (Cathartes aura). The final specimen was a distal tibiotarsus fragment identified to Golden Eagle (Aquila chrysaetos). This specimen was taken to the Museum of Southwestern Biology’s Division of Birds and identified with their skeletal comparative collection. It was distinguished from Bald Eagle (Haliaeetus leucocephalus) based off the morphology of the supratendinal bridge, which is more convex and more proximally robust in Golden Eagles compared to Bald Eagles. Eagles are acutely significant in Pueblo culture and their feathers are extremely valued (Beaglehole 1936; LaZar and Dombrosky 2022; Tyler 1991).
3.2.6 Passeriformes (n = 3)
Three passeriform specimens have been identified so far in the Haynie assemblage: one small fragmented humerus generally identified to the order-level, a tibia fragment identified to Corvidae (the family comprising jays and crows), and one large carpometacarpus identified as Raven (Corvus corax).
3.2.7 Columbiformes (n = 1)
Pigeons and doves are in the order Columbiformes, and one ulna fragment was recovered from Haynie. This specimen is most likely a Mourning Dove (Zenaida macroura). An order-level identification was used because the Crow Canyon comparative collection does not include a Band-tailed Pigeon (Columba fasciata), which is the only other member in the order Columbiformes to consider in the region.
3.3 Actinopterygii (n = 5)
This taxonomic class includes the ray-finned fishes. Three specimens have been identified to this general class: two ribs and one fragmented vertebra centrum. Following Nelson (2006, 35), fish specimens should no longer be referred to as pisces, as it is an antiquated taxonomic term. Similarly, for fishes of inland North America, the use of osteichthyes should no longer be used (Nelson 2006, 83). This is so for two interrelated reasons. First, this term has been replaced by the Euteleostoma designation. It successfully describes a monophyletic clade that includes tetrapods. Secondly, since Euteloestoma includes tetrapods, it includes lobe-finned fishes (Sarcopterygii). Lobe-finned fishes—like the coelacanth (Actinistia)—are not native fishes in inland North America during the late Holocene (Cloutier and Forey 1991). The use of osteichthyes should be accordingly abandoned. Instead, Actinoptergygii (ray-finned fishes) should be used because it is a more accurate class-level designation for archaeofaunas from the U.S. Southwest/Mexican Northwest.
3.3.1 Cypriniformes (n = 2)
This order includes carps, minnows, and suckers, which are common fishes in the aridland streams of the U.S. Southwest (Minckley and Marsh 2009; Sublette et al. 1990). These specimens are small intact vertebra. The lateral ridge morphology of centra can be used to identify vertebrae of fishes from U.S. Southwestern archaeofaunas to the order-level. Common orders of fishes found in rivers in the U.S. Southwest include Cypriniformes, Siluriformes, Lepisosteiformes, and Salmoniformes, and each of these orders have distinct vertebral morphology. These specimens are also notably small. It is possible inhabitants of the Haynie site used non-targeted methods to capture fishes, such as seining (Dombrosky et al. 2022). A focus on fishing practices offers a basic way to understand aquatic habitat use associated with the Simon Draw watershed.
3.4 Oviparous Animal (n = 1)
Eggshell specimens were identified as oviparous animal. These specimens are likely from Turkey, but a general identification was given because a scanning electron microscope could not be used to assess mammillary cone morphology (Beacham and Durand 2007; Conrad et al. 2016; Lapham et al. 2016). It is possible, though unlikely, that this eggshell could be from a lizard.
4 Taphonomy
Taphonomy is the study of how the material remains of organisms transition from the living world, to the lithosphere, and how they are subsequently recovered and studied by researchers (Lyman 1994). The most pressing taphonomic question for the Haynie archaeofaunal assemblage is: why are there so many unidentifiable specimens?
There are many interrelated factors that could lead to low identifiability, recall that the current identifiability rate at the Haynie site is 27.22%. The typical identifiability rate is 40–60% from previous Crow Canyon projects (Rawlings and Driver 2015; Driver et al. 1999). One of the most important taphonomic processes to consider here is fragmentation. Specimens retain fewer morphologically distinct features when an assemblage is highly fragmented (Cannon 2013). Humans are a prime taphonomic agent responsible for low identifiability considering that access to within-bone nutrients is facilitated by fragmentation (Wolverton 2002). Another agent to consider is poor bone preservation, which is caused by the degradation of organic and inorganic tissue. Poor preservation leads to brittle bone that crumbles when removed from its archaeological context, often leaving a distinctly bright breakage surface known as excavation damage. Many variables can contribute to poor bone preservation, and they include bone weathering, soil acidity, or the age of the archaeological deposit itself. Another critical taphonomic agent to consider are archaeofaunal analysts themselves. Analysts can influence identifiability by the amount of experience they have or by the identification protocols they follow (Gobalet 2001). Disentangling how these taphonomic agents influence an assemblage is difficult, but many multivariate methods are specifically designed to systematically sort through just such a morass of data.
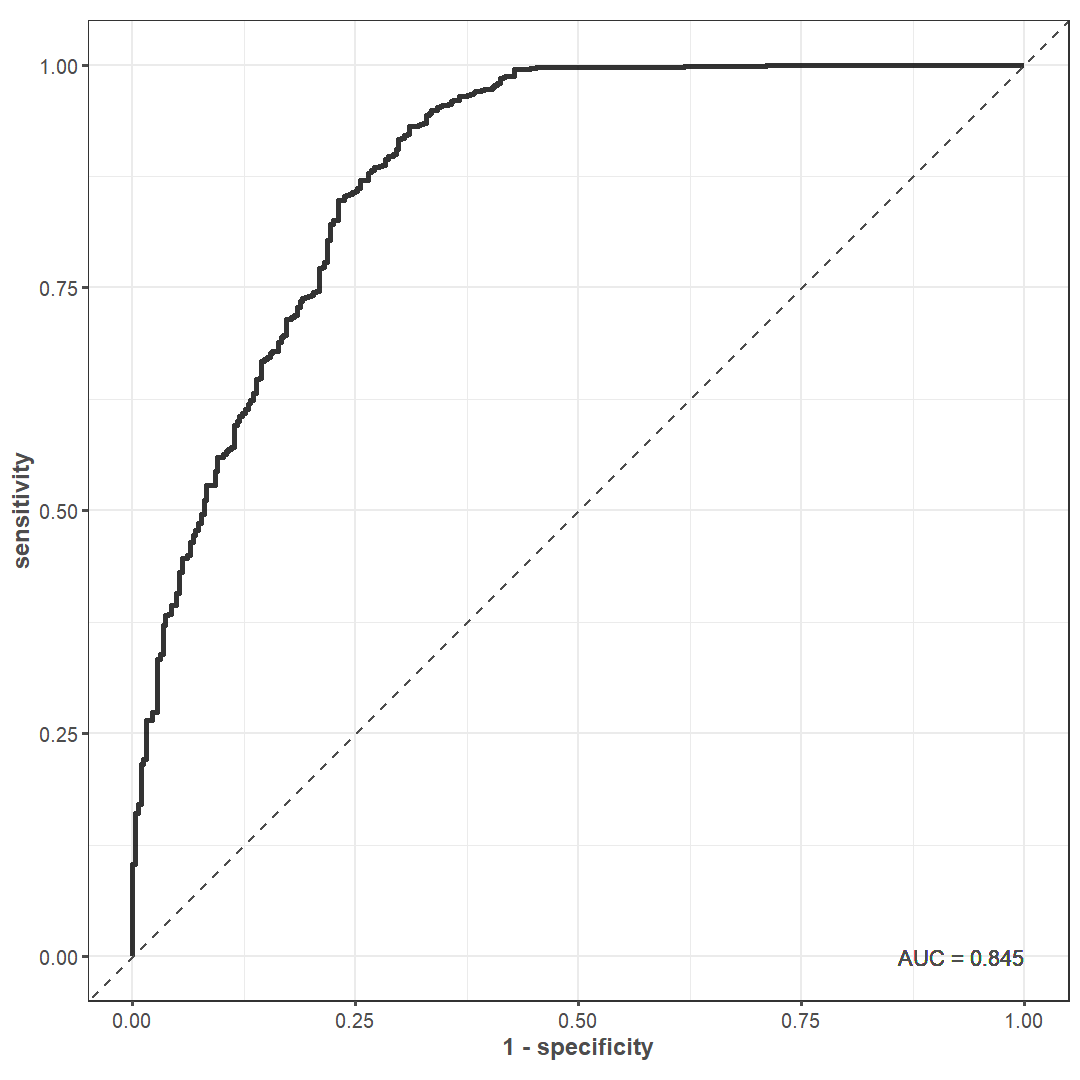
Predictive modeling is an effective tool in these circumstances. If a model can successfully predict unidentifiable specimens based on many different taphonomic variables, then an analyst can evaluate why the model behaves the way it does while accounting for every variable at once. The most important variables would drive unidentifiability in this scenario. Here, we use logistic regression to help achieve this goal. We supplied our logistic regression model with 16 predictor variables for every specimen: if it had 1) thick cortical bone, 2) excavation damage, 3) carnivore damage, 4) at least one intact end, 5) a spiral fracture, 6) a transverse fracture, 7) an irregular break, if it was 8) a shaft fragment, 9) made into an artifact, 10) eroded, 11) gnawed by rodents, 12) splintered, 13) root etched, 14) burned, 15) who the analyst was, and 16) its maximum length.
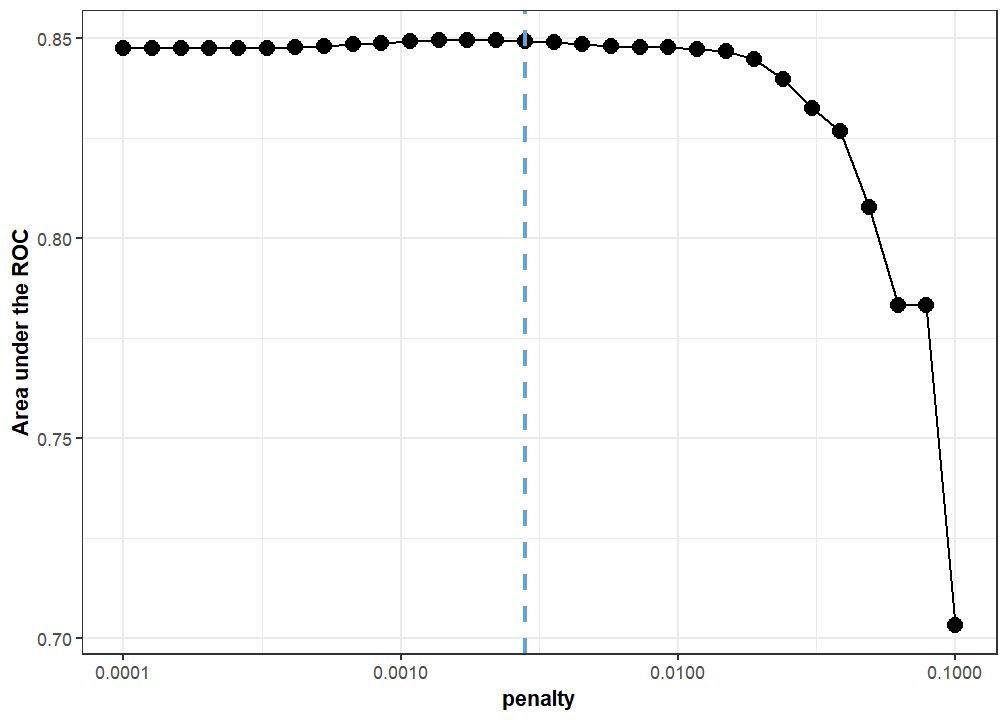
We included a penalty term in the model to safeguard against highly correlated predictor variables. We used grid search on a validation set to tune 30 candidate penalty values, and we picked the one with the highest area under the Receiver Operator Curve (ROC) for our final model (Figure 14). This model was 86.44% accurate on the training set, which is generally good. Interestingly, there was a large discrepancy between two further accuracy metrics. The Matthews correlation coefficient for the trained model is 0.64 while its F1 metric is 0.91. The F1 metric is markedly higher—indicating a model with high performance—because it describes how well the model predicts an event of interest. In this case, the event of interest is how well the model predicts unidentifiable specimens rather than identifiable ones (Figure 15; Figure 16). As such, this model fulfills the purpose of the current analysis.
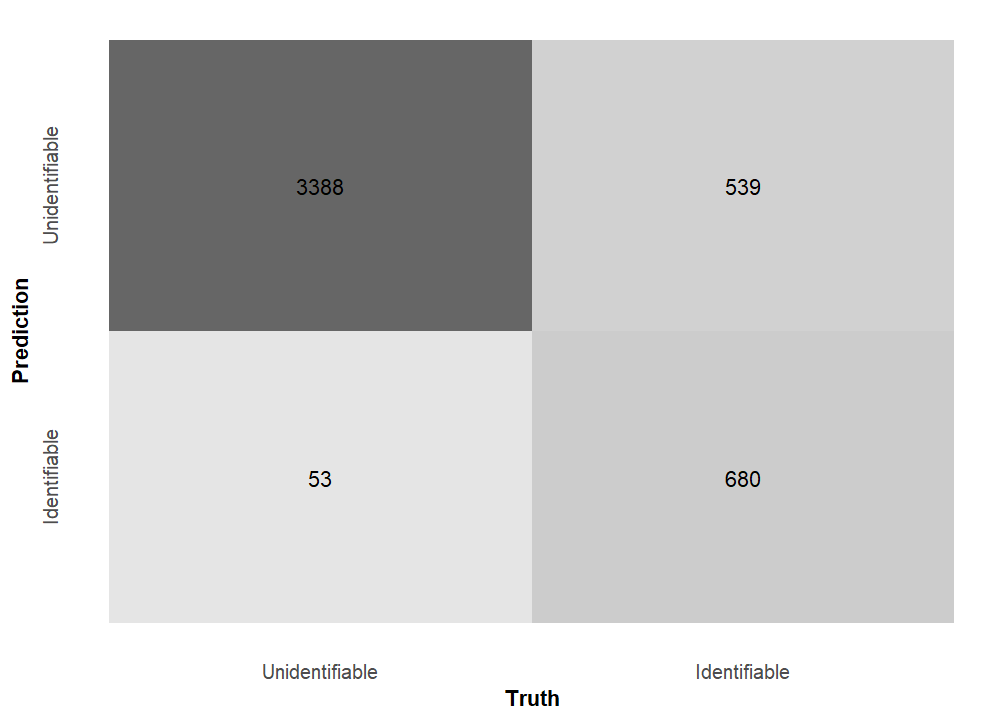
There are two main variables that explain why there are so many unidentifiable specimens at the Haynie site (Figure 17). The most important variable is whether or not a specimen has at least one intact end. This variable detracts from unidentifiability. In other words, the more intact a specimen is the more identifiable it is. This result indicates that the analysts and the identification protocol have a heavy influence on this data, but they do so in a way that enhances data quality. An assemblage with high identifiability on specimens lacking morphologically distinct features (i.e., intact ends) would be a red flag. The most important variable contributing to unidentifiability—the main variable of interest for the purposes here—is whether or not the specimen has thick cortical bone. This results suggests that specimens lacking intact portions but possessing thick cortical bone are overwhelmingly unidentifiable. Thus, the main taphonomic agents responsible for low identifiability are likely the inhabitants of the Haynie site.
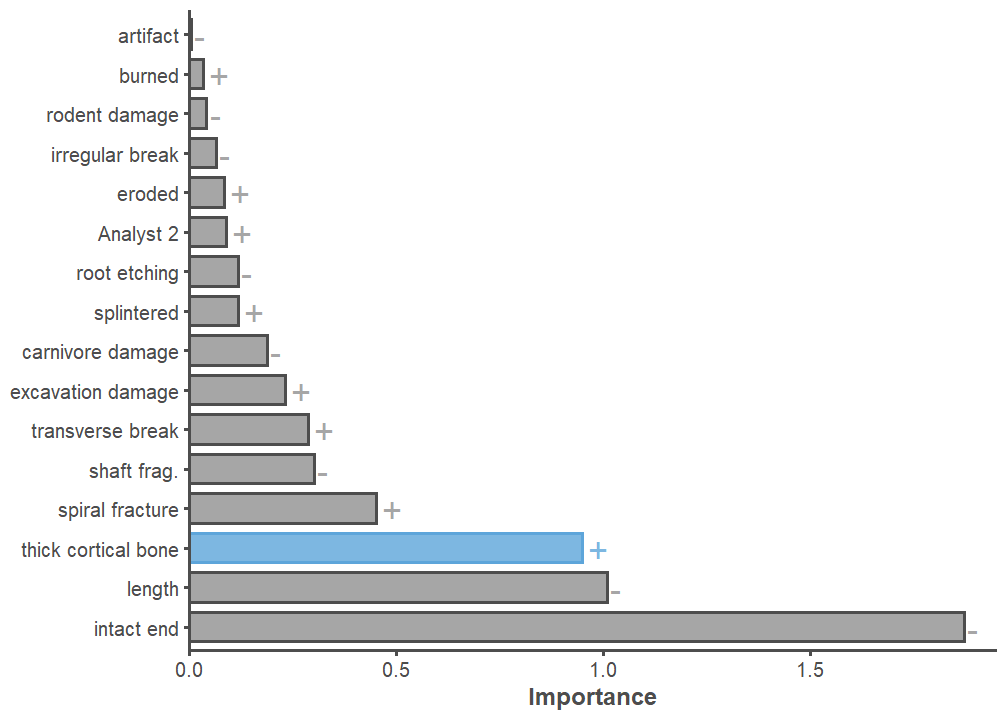
Medium-to-large mammal fragmentation is a key taphonomic feature of this assemblage, and will be vital to incorporate into future zooarchaeological work. Evaluating the extent and intensity of fragmentation per skeletal element of medium and large mammals is of particular interest (sensu Wolverton 2002). The presence of many unidentifiable specimens with thick cortical bone could be related to many medium and large mammals at the site. In this scenario, these specimens could indicate efficient foraging practices, where Haynie hunters were able to consistently acquire high-ranked prey. Another possible interpretation is that foraging efficiency was low and that large prey were intensively and extensively exploited for nutrients. The calculation of fragmentation rates per skeletal part and across the skeleton of individual artiodactyls will help resolve this issue. For either of these scenarios to be true, the current somewhat moderate Artiodactyl Index (0.34) would have to be a sampling anomaly. Another critical factor to add to future models is whether specimens come from intact or disturbed deposits, as the Haynie site does include clear looter’s pits and areas of mechanical disturbance (Throgmorton et al. 2022).
5 Taxonomic Diversity and Representative Sampling
The Lakeview Community is located in a shallow, wide valley that is part of the Simon Draw watershed. Simon Draw itself flowed close to the site (Throgmorton et al. 2022). As such, the Lakeview Community was likely located close to marshy habitat in the past. A natural corridor like this could have facilitated the movement of both birds and large game, supplying highly diverse wild resources to the Haynie site residents. Here, we test this hypothesis by modeling identification accumulation rates and evenness. We compare these estimations to three other sites in the northern U.S. Southwest and control for sample size effects using rarefaction. This analysis serves multiple purposes at once. Not only can we accurately gauge and compare taxonomic diversity across sites, we may also assess whether sampling efforts have been sufficient enough to provide accurate taxonomic representation.
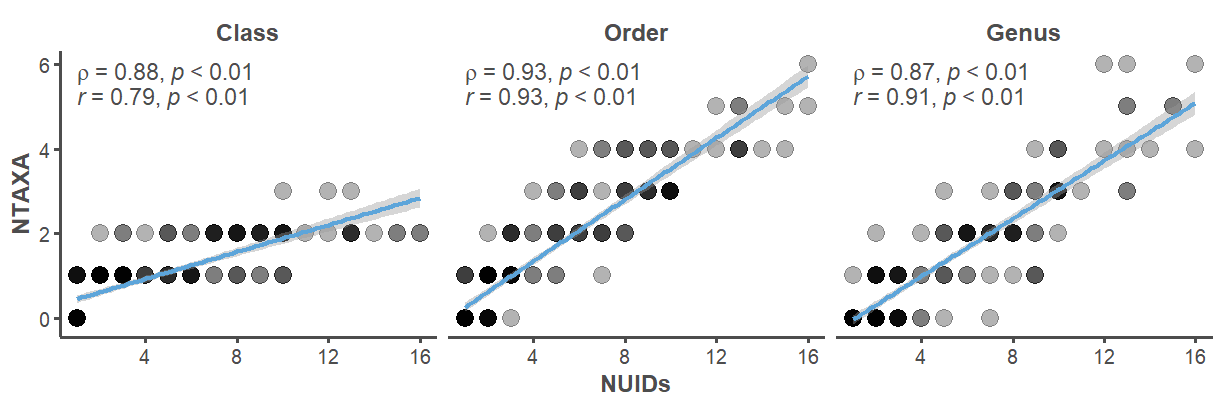
This analysis relies on a non-standard quantitative unit to estimate taxonomic richness called the Number of Unique Identifications (NUIDs). It is a tally of the different identification types present in a specific context, meaning it can include standard taxonomic identifications (e.g., Odocoileus sp.) and non-standard identifications (e.g., medium artiodactyl). This unit serves as a proxy for taxonomic richness to help gauge patterns in sampling and recovery. We prefer this unit over the common Number of Taxa (NTAXA) for three reasons: 1) it is simpler to calculate when dealing with large mixed assemblages identified to a variety of taxonomic levels, 2) it does not require the selection of an arbitrary taxonomic group from which to aggregate all lower units within, and 3) it is strongly correlated with NTAXA when calculated at multiple levels of taxonomic resolution (Figure 18).
We compared how the number of new identifications accumulate as NISP increases at Haynie to three other archaeofaunal assemblages: Sand Canyon Pueblo (5MT765), Ponsipa’akeri (LA 297), and Shields Pueblo (Figure 19). The Haynie site is not particularly diverse compared to other assemblages in the northern U.S. Southwest. Taxonomic richness at Haynie is similar to terminal Pueblo III sites in the same region (Sand Canyon) or even Pueblo IV sites from the Northern Rio Grande (Ponsipa’akeri). Shields Pueblo, a nearby contemporary Great House site, has lower than expected diversity. The distribution of Haynie taxonomic abundance is unlike Great House sites in the same region, but it is like later sites (Figure 20). Perhaps control of wild resources and foodways is leading to lower-than-expected diversity measures in such a potentially diverse environmental setting. Additionally, the duration of site occupation could impact accumulation rates (sensu Varien and Potter 1997; Varien and Mills 1997; Varien and Ortman 2005). Controlling for site occupation and increasing sample size will clarify these patterns.
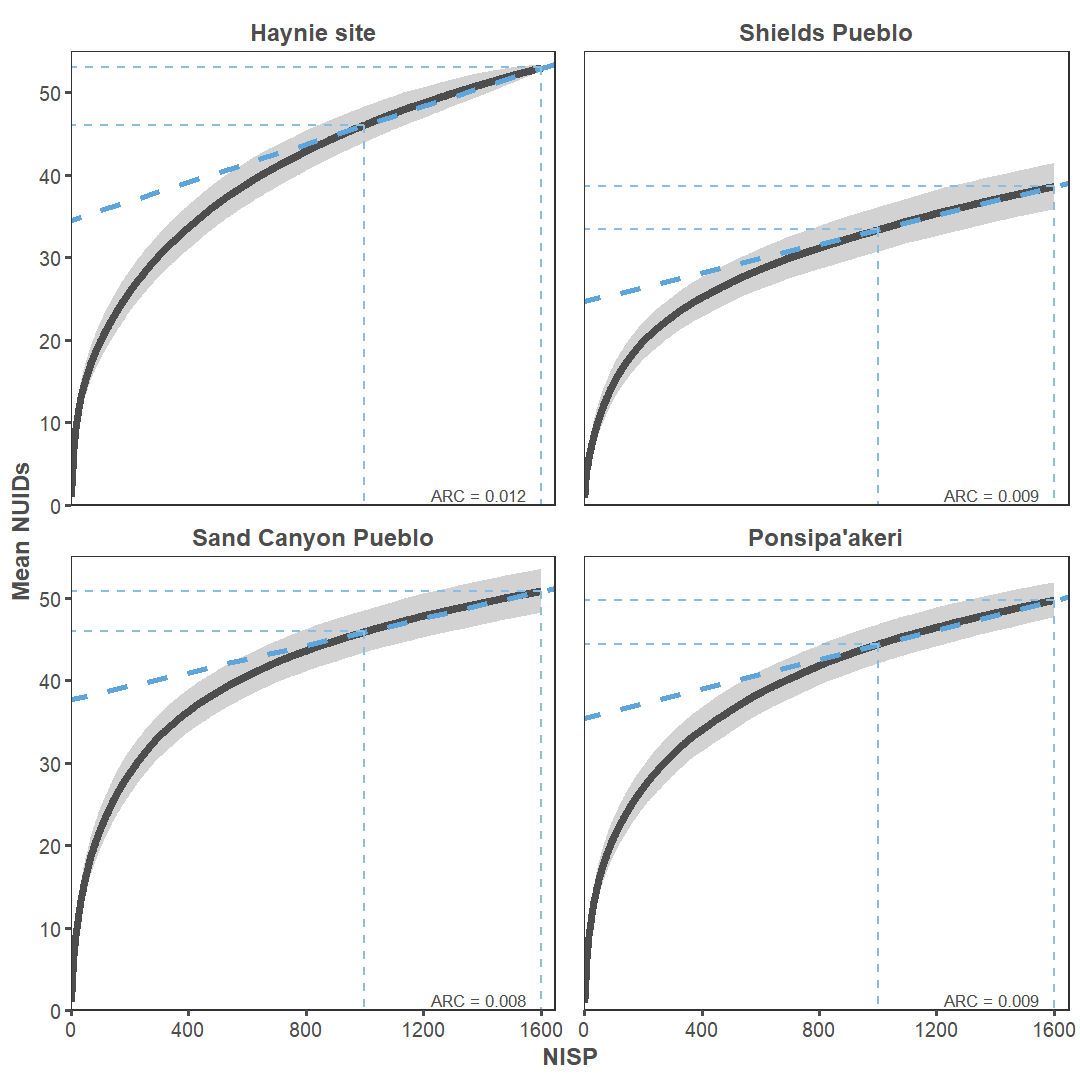
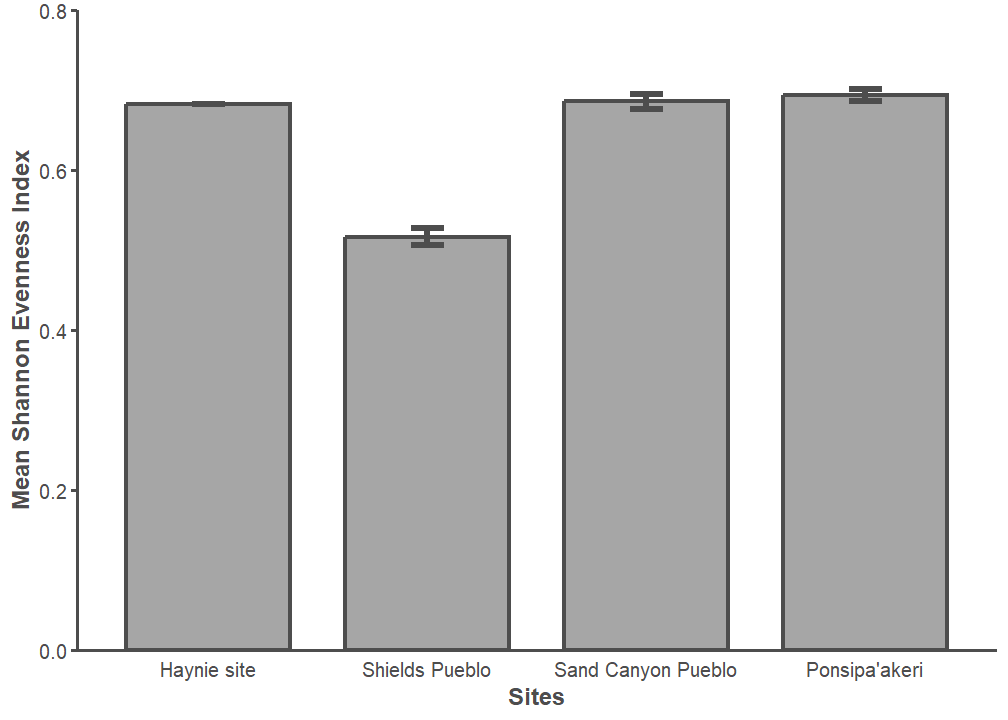
Increasing sample size is crucial, because our sampling effort is not yet sufficient enough to provide accurate taxonomic representation. The Average Rate of Change (ARC) is high at each site when the sample is beyond an NISP of 1,000, but it is highest at Haynie. A new identification is added every 111 to 125 identified specimens at the other sites, and a new identification type is added for every 83 specimens at Haynie. We consider 83 identified specimens low sampling effort for every new identification type gained. Any final word on Haynie taxonomic diversity must wait until the accumulation of new identifications levels-off as sampling intensity increases.
6 Conclusion
We have demonstrated that data produced during the first year of archaeofaunal analysis at the Haynie site is high quality, and it points to three specific areas of future research. First, artiodactyl exploitation could be a significant component of the Haynie archaeofaunal assemblage. The ratio of mule deer to pronghorn suggests that local hunting practices dominated, which is interesting to think about when contemplating how bison were procured at the site. The local or non-local procurement of bison will be important for understanding the economic structure of the Lakeview Community, while also helping to reconstruct the historical ecology of bison on the Colorado Plateau. The taphonomic analysis also suggested that artiodactyl processing is a prime reason there are so many unidentifiable bones, but predictors related to disturbance context need to be added to the taphonomic model. Second, the presence of an intact wolf mandible—from an advanced age class—points to a number of future questions about resource procurement and the nature of human-carnivore interactions: was the mandible intentionally placed, was the individual cared for in anyway, and was the wolf local to the site? Finally, the analysis of taxonomic diversity showed that the Haynie archaeofaunal assemblage is not significantly diverse compared to other sites, but that more data is needed to acquire a representative picture of taxonomic composition. Zooarchaeological material is poised to answer a number of questions relevant to the Northern Chaco Outliers Project and to the archaeology of human-environment interaction in the U.S. Southwest and beyond.
7 Acknowledgements
Thank you to the Museum of Southwestern Biology’s Division of Mammals and Division of Birds, with specific regard to Adrienne Raniszewski, Jonathan Dunnum, Joseph Cook, and Andy Johnson. Doug Smith and Erin Stahler, of the Yellowstone Wolf Project, kindly provided gray wolf mortality data. Personal thanks to Lee Lyman, Martin Welker, and Steve Wolverton for stimulating zooarchaeological conversation and advice.
8 References
Adams, Bradley J., and Pam J. Crabtree. 2012. Comparative Osteology a Laboratory and Field Guide of Common North American Animals. Boston, MA: Elsevier/Academic Press.
Andrews, Peter, and Jill Cook. 1990. Owls, Caves and Fossils: Predation, Preservation and Accumulation of Small Mammal Bones in Caves, with an Analysis of the Pleistocene Cave Faunas from Westbury-Sub-Mendip, Somerset, UK. Chicago, IL: University of Chicago Press.
Badenhorst, Shaw, and Jonathan C. Driver. 2009. “Faunal Changes in Farming Communities from Basketmaker II to Pueblo III (A.D. 1-1300) in the San Juan Basin of the American Southwest.” Journal of Archaeological Science 36 (9): 1832–41. https://doi.org/10.1016/j.jas.2009.04.006.
———. 2015. “Faunal Remains.” In The Archaeology of Albert Porter Pueblo (Site 5MT123): Excavations at a Great House Community Center in Southwestern Colorado, edited by Susan C. Ryan, 448–75. Cortez, CO: Crow Canyon Archaeological Center.
Badenhorst, Shaw, Jonathan C. Driver, and Susan C. Ryan. 2019. “Desirable Meat: The Social Context of Meat Procurement at Albert Porter Pueblo, a Great House Community in the Central Mesa Verde Region.” KIVA 85 (3): 191–213. https://doi.org/10.1080/00231940.2019.1579442.
Badenhorst, Shaw, Jonathan C. Driver, and Steve Wolverton. In press. “The Consumption of Small Rodents in the Ancestral Puebloan World of the American Southwest.” In Crow Canyon Archaeological Center 40th Anniversary Volume, edited by Susan C. Ryan. Salt Lake City: The University of Utah Press.
Baker, Brenda J., Dupras Tosha L., and Matthew W. Tocheri. 2005. The Osteology of Infants and Children. Vol. 12. College Station, TX: Texas A&M University Press.
Baker, Bruce W., and Edward P. Hill. 2003. “Beaver (Castor Canadensis).” In Wild Mammals of North America: Biology, Management, and Conservation, edited by George A. Feldhamer, Bruce C. Thompson, and Joseph A. Chapman, 2nd ed., 288–310. Baltimore, MD: The Johns Hopkins University Press.
Beacham, E. Bradley, and Stephen R. Durand. 2007. “Eggshell and the Archaeological Record: New Insights into Turkey Husbandry in the American Southwest.” Journal of Archaeological Science 34 (10): 1610–21. https://doi.org/10.1016/j.jas.2006.11.015.
Beaglehole, Ernest. 1936. Hopi Hunting and Ritual. Yale University Publications in Anthropology 4. New Haven, CT: Yale University Press.
Binford, Lewis R. 1978. Nunamiut Ethnoarchaeology. New York: Academic Press.
Bocinsky, R. Kyle, Jason A. Cowan, Timothy A. Kohler, and C. David Johnson. 2012. “How Hunting Changes the VEP World, and How the VEP World Changes Hunting.” In Emergence and Collapse of Early Villages: Models of Central Mesa Verde Archaeology, edited by Timothy A. Kohler and Mark Varien, 145–52. Berkeley: University of California Press.
Broughton, Jack M., Michael D. Cannon, Frank E. Bayham, and David A. Byers. 2011. “Prey Body Size and Ranking in Zooarchaeology: Theory, Empirical Evidence, and Applications from the Northern Great Basin.” American Antiquity 76 (3): 403–28. https://doi.org/10.7183/0002-7316.76.3.403.
Broughton, Jack M., and Shawn D. Miller. 2016. Zooarchaeology and Field Ecology: A Photographic Atlas. Salt Lake City: The University of Utah Press.
Brown, Donald Nelson. 1967. “The Distribution of Sound Instruments in the Prehistoric Southwestern United States.” Ethnomusicology 11 (1): 71–90. https://doi.org/10.2307/850499.
Burger, Rachel. 2021. “Food Security in Ancestral Tewa Coalescent Communities: The Zooarchaeology of Sapa’owingeh in the Northern Rio Grande, New Mexico.” Doctoral Dissertation, Dallas, TX: Department of Anthropology, Southern Methodist University. https://scholar.smu.edu/hum_sci_anthropology_etds/13/.
Cannon, Michael D. 2013. “NISP, Bone Fragmentation, and the Measurement of Taxonomic Abundance.” Journal of Archaeological Method and Theory 20 (3): 397–419. https://doi.org/10.1007/s10816-012-9166-z.
Cates, Kari M. (Schmidt). 2020. “Faunal Remains from Greenstone Pueblo (5MT6970).” Manuscript on file Crow Canyon Archaeological Center, Cortez, CO.
Chavez, Angela Marie. 2008. “Comparative Vertebral Morphology in Medium-Sized North American Artiodactyla.” The Artifact 46: 1–17.
Cloutier, Richard, and Peter L. Forey. 1991. “Diversity of Extinct and Living Actinistian Fishes (Sarcopterygii).” In The Biology of Latimeria Chalumnae and Evolution of Coelacanths, edited by John A. Musick, Michael N. Bruton, and Eugene K. Balon, 59–74. Springer.
Cohen, Alan, and D. Serjeantson. 1996. A Manual for the Identification of Bird Bones from Archaeological Sites. London: Archetype Publications.
Conrad, Cyler, Emily Lena Jones, Seth D. Newsome, and Douglas W. Schwartz. 2016. “One Isotopes, Eggshell and Turkey Husbandry at Arroyo Hondo Pueblo.” Journal of Archaeological Science: Reports 10: 566–74. https://doi.org/10.1016/j.jasrep.2016.06.016.
Dombrosky, Jonathan, Thomas F. Turner, Alexandra Harris, and Emily Lena Jones. 2022. “Body Size from Unconventional Specimens: A 3D Geometric Morphometrics Approach to Fishes from Ancestral Pueblo Contexts.” Journal of Archaeological Science 142: 105600. https://doi.org/10.1016/j.jas.2022.105600.
Driver, Jonathan C. 1992. “Identification, Classification and Zooarchaeology.” Circaea 9 (1): 35–47.
———. 2002. “Faunal Variation and Change in the Northern San Juan Region.” In Seeking the Center Place: Archaeology and Ancient Communities in the Mesa Verde Region, edited by Mark D. Varien and Richard H. Wilshusen, 143–60. Salt Lake City: The Univeristy of Utah Press.
———. 2006. Crow Canyon Archaeological Center Manual for Description of Vertebrate Remains. 8th ed. Cortez, CO: Crow Canyon Archaeological Center.
———. 2011. “Human Impacts on Animal Populations in the American Southwest.” In Movement, Connectivity and Landscape Change in the Ancient Southwest, edited by Margaret C. Nelson and Colleen Strawhacker, 179–98. Boulder: University Press of Colorado.
———. 2011. “Identification, Classification and Zooarchaeology (Featured Reprint and Invited Comments).” Ethnobiology Letters 2: 19–39. https://doi.org/10.14237/ebl.2.2011.32.
Driver, Jonathan C., Michael J. Brand, Lianne Lester, and Natalie D. Munro. 1999. “Faunal Studies.” In The Sand Canyon Archaeological Project: Site Testing, edited by Mark D. Varien. Cortez, CO: Crow Canyon Archaeological Center. https://www.crowcanyon.org/ResearchReports/SiteTesting/Text/Report.asp.
Driver, Jonathan C., and Joshua R. Woiderski. 2008. “Interpretation of the "Lagomorph Index" in the American Southwest.” Quaternary International 185: 3–11. https://doi.org/10.1016/j.quaint.2007.09.022.
Elbroch, Mark. 2006. Animal Skulls: A Guide to North American Species. Mechanicsburg, PA: Stackpole Books.
Ellyson, Laura. 2014. “Resource Intensification of Small Game Use at Goodman Point, Southwestern Colorado.” Master's Thesis, Denton: Department of Geography, University of North Texas. https://digital.library.unt.edu/ark:/67531/metadc699883/.
Fernández-Jalvo, Yolanda, and Peter Andrews. 2016. Atlas of Taphonomic Identifications: 1001+ Images of Fossil and Recent Mammal Bone Modification. Springer.
France, Diane L. 2009. Human and Nonhuman Bone Identification: A Color Atlas. Boca Raton, FL: CRC Press.
Frisbie, Theodore R. 1967. “The Excavation and Interpretation of the Artificial Leg Basket Maker III-Pueblo I Sites near Corrales, New Mexico.” Master's Thesis, Albuquerque: Department of Anthropology, University of New Mexico.
Gilbert, B. Miles. 1980. Mammalian Osteology. Laramie, WY: B. M. Gilbert.
Gilbert, B. Miles, Larry D. Martin, and Howard G. Savage. 1981. Avian Osteology. Laramie, WY: B. M. Gilbert.
Gipson, Philip S., Warren B. Ballard, Ronald M. Nowak, and L. David Mech. 2000. “Accuracy and Precision of Estimating Age of Gray Wolves by Tooth Wear.” The Journal of Wildlife Management 64 (3): 752–58. https://doi.org/10.2307/3802745.
Gobalet, Kenneth W. 2001. “A Critique of Faunal Analysis: Inconsistency Among Experts in Blind Tests.” Journal of Archaeological Science 28 (4): 377–86. https://doi.org/10.1006/jasc.2000.0564.
Grayson, Donald K. 1978. “Minimum Numbers and Sample Size in Vertebrate Faunal Analysis.” American Antiquity 43 (1): 53–65. https://doi.org/10.2307/279631.
———. 1981. “The Effects of Sample Size on Some Derived Measures in Vertebrate Faunal Analysis.” Journal of Archaeological Science 8 (1): 77–88. https://doi.org/10.1016/0305-4403(81)90013-3.
———. 1984. Quantitative Zooarchaeology: Topics in the Analysis of Archaeological Faunas. Orlando, FL: Academic Press. https://www.sciencedirect.com/book/9780122972805/quantitative-zooarchaeology.
Greenwell, Brandon M., and Bradley C. Boehmke. 2020. “Variable Importance Plots—An Introduction to the vip Package.” The R Journal 12 (1): 343–66. https://doi.org/10.32614/RJ-2020-013.
Guiry, Eric, Trevor J. Orchard, Suzanne Needs-Howarth, and Paul Szpak. 2021. “Isotopic Evidence for Garden Hunting and Resource Depression in the Late Woodland of Northeastern North America.” American Antiquity 86 (1): 90–110. https://doi.org/10.1017/aaq.2020.86.
Hargrave, Lyndon L., and Steven D. Emslie. 1979. “Osteological Identification of Sandhill Crane Versus Turkey.” American Antiquity 44 (2): 295–99. https://doi.org/10.2307/279079.
Hastie, Trevor, Robert Tibshirani, and Jerome H. Friedman. 2009. The Elements of Statistical Learning: Data Mining, Inference, and Prediction. Springer.
Hayes, Alden C., and James A. Lancaster. 1975. Badger House Community, Mesa Verde National Park. Wetherill Mesa Studies, Publication in Archeology 7E. Washington D.C.: U.S. Department of the Interior, National Park Service.
Hill, Erica. 2000. “The Contextual Analysis of Animal Interments and Ritual Practice in Southwestern North America.” KIVA 65 (4): 361–98. https://doi.org/10.1080/00231940.2000.11758417.
Hillson, Simon. 1986. Teeth. New York: Cambridge University Press.
———. 1996. Mammal Bones and Teeth: An Introductory Guide to Methods of Identification. London: Institute of Archaeology, University College London.
Jacobson, Jodi A. 2003. “Identification of Mule Deer (Odocoileus Hemionus) and White-Tailed Deer (Odocoileus Virginianus) Postcranial Remains as a Means of Determining Human Subsistence Strategies.” Plains Anthropologist 48 (187): 287–97. https://doi.org/10.1080/2052546.2003.11949269.
James, Gareth, Daniela Witten, Trevor Hastie, and Robert Tibshirani. 2013. An Introduction to Statistical Learning: With Applications in R. Springer.
Kachigan, Sam Kash. 1991. Multivariate Statistical Analysis: A Conceptual Introduction. 2nd ed. New York: Radius Press.
Kuhn, Max, and Kjell Johnson. 2013. Applied Predictive Modeling. Springer.
Kuhn, Max, and Julia Silge. 2022. Tidy Modeling with R. Sebastopol, CA: O’Reilly Media, Inc. https://www.tmwr.org/.
Ladd, Edmund J. 1963. “Zuni Ethno-Ornithology.” Master's Thesis, Albuquerque: Department of Anthropology, University of New Mexico.
Lang, Richard W., and Arthur H Harris. 1984. The Faunal Remains from Arroyo Hondo Pueblo, New Mexico: A Study in Short-Term Subsistence Change. Arroyo Hondo Archaeological Series. Santa Fe, NM: School of American Research Press.
Lapham, Heather A., Gary M. Feinman, and Linda M. Nicholas. 2016. “Turkey Husbandry and Use in Oaxaca, Mexico: A Contextual Study of Turkey Remains and SEM Analysis of Eggshell from the Mitla Fortress.” Journal of Archaeological Science: Reports 10: 534–46. https://doi.org/10.1016/j.jasrep.2016.05.058.
LaZar, Miranda, and Jonathan Dombrosky. 2022. “Exploring Raptor Exchange.” Archaeology Southwest Magazine 35 (1 & 2): 29–31. https://www.archaeologysouthwest.org/product/asw35-1-2/.
Lyman, R. Lee. 1994. Vertebrate Taphonomy. Cambridge: Cambridge University Press.
———. 2002. “Taxonomic Identification of Zooarchaeological Remains.” The Review of Archaeology 23 (2): 13–20.
———. 2008. Quantitative Paleozoology. Cambridge: Cambridge University Press.
———. 2019. “Assumptions and Protocol of the Taxonomic Identification of Faunal Remains in Zooarchaeology: A North American Perspective.” Journal of Archaeological Method and Theory 26 (4): 1376–1438. https://doi.org/10.1007/s10816-019-09414-0.
Martin, Jeff M., Rachel A. Martin, and Jim I. Mead. 2017. “Late Pleistocene and Holocene Bison of the Colorado Plateau.” The Southwestern Naturalist 62 (1): 14–28. https://doi.org/10.1894/0038-4909-62.1.14.
Minckley, Wendell L., and Paul C. Marsh. 2009. Inland Fishes of the Greater Southwest: Chronicle of a Vanishing Biota. Tucson: The University of Arizona Press.
Monagle, Victoria, and Emily Lena Jones. 2020. “Dog Life and Death in an Ancestral Pueblo Landscape.” In Dogs: Archaeology beyond Domestication, edited by Brandi Bethke and Amanda Burtt, 45–71. Gainesville: University Press of Florida.
Muir, Robert J., and Jonathan C. Driver. 2002. “Scale of Analysis and Zooarchaeological Interpretation: Pueblo III Faunal Variation in the Northern San Juan Region.” Journal of Anthropological Archaeology 21 (2): 165–99. https://doi.org/10.1006/jaar.2001.0392.
———. 2004. “Identifying Ritual Use of Animals in the Northern American Southwest.” In Behaviour Behind Bones: The Zooarchaeology of Ritual, Religion, Status, and Identity, edited by Sharyn Jones O’Day, Wim Van Neer, and Anton Ervynck, 128–43. Oxford: Oxbow Books.
Nelson, Joseph S. 2006. Fishes of the World. Hoboken, NJ: John Wiley.
Olsen, Sandra L. 1979. “A Study of Bone Artifacts from Grasshopper Pueblo, AZ P:14:1.” KIVA 44 (4): 341–73. https://doi.org/10.1080/00231940.1979.11757924.
———. 1980. “Bone Artifacts from Kinishba Ruin: Their Manufacture and Use.” KIVA 46 (1-2): 39–67. https://doi.org/10.1080/00231940.1980.11757947.
Olsen, Stanley John. 1964. Mammal Remains from Archaeological Sites. Cambridge, MA: Peabody Museum.
Payne, Richard W. 1991. “Bone Flutes of the Anasazi.” KIVA 56 (2): 165–77. https://doi.org/10.2307/30247265.
Potter, James M. 2000. “Pots, Parties, and Politics: Communal Feasting in the American Southwest.” American Antiquity 65 (3): 471–92. https://doi.org/10.2307/2694531.
———. 2004. “Hunting and Social Differentiation in the Late Prehispanic American Southwest.” In Behaviour Behind Bones: Zooarchaeology of Ritual, Religion, Status and Identity, edited by Sharyn Jones O’Day, Wim Van Neer, and Anton Ervynck, 285–92. Oxford: Oxbow Books.
Potter, James M., and Scott G. Ortman. 2004. “Community and Cuisine in the Prehispanic American Southwest.” In Identity, Feasting, and the Archaeology of the Greater Southwest, edited by Barbara J. Mills, 173–91. Boulder: University Press of Colorado.
R Core Team. 2022. R: A Language and Environment for Statistical Computing. 4.2.0 ed. Vienna, Austria: R Foundation for Statistical Computing. https://www.r-project.org/.
Rawlings, Tiffany A. 2006. “Faunal Analysis and Meat Procurement: Reconstructing the Sexual Division of Labor at Shields Pueblo, Colorado.” Doctoral Dissertation, Burnaby, BC: Department of Archaeology, Simon Fraser University. https://summit.sfu.ca/item/2374.
Rawlings, Tiffany A., and Jonathan C. Driver. 2015. “Faunal Remains from Shields Pueblo.” In The Archaeology of Shields Pueblo (Site 5MT3807): Excavations at a Mesa-Top Community Center in Southwestern Colorado, edited by Susan C. Ryan, 186–221. Cortez, CO: Crow Canyon Archaeological Center.
Roze, Uldis, and Linda M. Ilse. 2003. “Porcupine (Erethizon Dorsatum).” In Wild Mammals of North America: Biology, Management, and Conservation, edited by George A. Feldhamer, Bruce C. Thompson, and Joseph A. Chapman, 2nd ed., 371–80. Baltimore, MD: The Johns Hopkins University Press.
Ryan, Susan C. 2016. “The Northern Chaco Outliers Project: A Proposal to Conduct Archaeological Testing at the Haynie Site, Southwestern Colorado.” Manuscript on file Crow Canyon Archaeological Center, Cortez, CO. Submitted to Colorado State Historic Preservation Office, Denver, CO.
Smart, Tamela S. 2009. “Carpals and Tarsals of Mule Deer, Black Bear and Human: An Osteology Guide for the Archaeologist.” Master's Thesis, Pullman: Department of Anthropology, Western Washington University. https://cedar.wwu.edu/cgi/viewcontent.cgi?referer=&httpsredir=1&article=1018&context=wwuet.
Spielmann, Katherine A., and Eric A. Angstadt-Leto. 1996. “Hunting, Gathering, and Health in the Prehistoric Southwest.” In Evolving Complexity and Environmental Risk in the Prehistoric Southwest, edited by Joseph A. Tainter, and Bonnie B. Tainter, 79–106. Reading, MA: CRC Press.
Strand, Jennifer G. 1998. “An Analysis of the Homol’ovi Fauna with Emphasis of Ritual Behavior.” Doctoral Dissertation, Tucson: Department of Anthropology, University of Arizona. https://repository.arizona.edu/handle/10150/288835.
Sublette, James E., Michael D. Hatch, and Mary F. Sublette. 1990. The Fishes of New Mexico. Albuquerque: University of New Mexico Press.
Throgmorton, Kellam, Susan C. Ryan, Benjamin Bellorado, Jeremy Grundvig, Steve Copeland, and Timothy Wilcox. 2022. “Excavation and Additional Studies at The Haynie Site (5MT1905) by the Crow Canyon Archaeological Center.” Annual Report 2021. Cortez, CO: Crow Canyon Archaeological Center. https://www.crowcanyon.org/wp-content/uploads/2022/02/ncop-annual-report-2021-final-1.pdf.
Twiss, Katheryn. 2007. The Archaeology of Food and Identity. Center for Archaeological Investigations, Occasional Paper 34. Carbondale: Southern Illinois University.
Tyler, Hamilton A. 1991. Pueblo Birds and Myths. Flagstaff, AZ: Northland Publishing Company.
Varien, Mark D., and Barbara J. Mills. 1997. “Accumulations Research: Problems and Prospects for Estimating Site Occupation Span.” Journal of Archaeological Method and Theory 4 (2): 141–91. https://doi.org/10.1007/BF02428057.
Varien, Mark D., and Scott G. Ortman. 2005. “Accumulations Research in the Southwest United States: Middle-Range Theory for Big-Picture Problems.” World Archaeology 37 (1): 132–55. https://doi.org/10.1080/0043824042000329603.
Varien, Mark D., and James M. Potter. 1997. “Unpacking the Discard Equation: Simulating the Accumulation of Artifacts in the Archaeological Record.” American Antiquity 62 (2): 194–213. https://doi.org/10.2307/282506.
von den Driesch, Angela. 1976. A Guide to the Measurement of Animal Bones from Archaeological Sites. Peabody Museum Press Bulletin 1. Cambridge, MA: Harvard University.
Watson, Adam S., and Matthew A. Gleason. 2016. “A Comparative Assessment of Texture Analysis Techniques Applied to Bone Tool Use-Wear.” Surface Topography: Metrology and Properties 4 (2): 024002. https://doi.org/10.1088/2051-672x/4/2/024002.
Wayne, Robert K., Eli Geffen, Derek J. Girman, Klaus P. Koepfli, Lisa M. Lau, and Charles R. Marshall. 1997. “Molecular Systematics of the Canidae.” Systematic Biology 46 (4): 622–53. https://doi.org/10.1093/sysbio/46.4.622.
Welker, Martin H., David A. Byers, and Sarah B. McClure. 2021. “‘I Wanna Be Your Dog’: Evaluating the Efficacy of Univariate and Multivariate Methods for Differentiating Domestic and Wild Canids in North America.” International Journal of Osteoarchaeology 31 (2): 196–206. https://doi.org/10.1002/oa.2939.
White, Tim D., Michael T. Black, and Pieter A. Folkens. 2012. Human Osteology. 3rd ed. Cambridge, MA: Academic Press.
Wickham, Hadley. 2010. “A Layered Grammar of Graphics.” Journal of Computational and Graphical Statistics 19 (1): 3–28. https://doi.org/10.1198/jcgs.2009.07098.
Wickham, Hadley, Mara Averick, Jennifer Bryan, Winston Chang, Lucy D’Agostino McGowan, Romain François, Garrett Grolemund, Alex Hayes, Lionel Henry, and Jim Hester. 2019. “Welcome to the Tidyverse.” Journal of Open Source Software 4 (43): 1686. https://doi.org/10.21105/joss.01686.
Wolverton, Steve. 2002. “NISP:MNE and %Whole in Analysis of Prehistoric Carcass Exploitation.” North American Archaeologist 23 (2): 85–100. https://doi.org/10.2190/egdq-cq1q-lld2-h3tp.
———. 2013. “Data Quality in Zooarchaeological Faunal Identification.” Journal of Archaeological Method and Theory 20 (3): 381–96. https://doi.org/10.1007/s10816-012-9161-4.
Wolverton, Steve, and Lisa Nagaoka. 2018. “Zooarcheology: Investigating Past Interactions Between Humans and Other Animals.” In Ethnozoology, edited by Rômulo Romeu Nóbrega Alves and Ulysses Paulino Albuquerque, 25–43. Cambridge, MA: Academic Press.
Citation
BibTeX citation:
@incollection{dombrosky2023,
author = {Dombrosky, Jonathan and Gilmore, Eric},
editor = {Throgmorton, Kellam and C. Ryan, Susan and Dombrosky,
Jonathan and Bellorado, Benjamin and Copeland, Steven and Hughes,
Kate and Merewether, Jamie},
title = {Year {One} of {Archaeofaunal} {Analysis} at the {Haynie}
{Site} {(5MT1905)}},
booktitle = {Excavation and Additional Studies at The Haynie Site
(5MT1905) by the Crow Canyon Archaeological Center Annual Report
2022},
pages = {45-88},
date = {2023},
url = {https://www.crowcanyon.org/wp-content/uploads/2023/02/ncop-annual-report-2022-final.pdf},
langid = {en}
}
For attribution, please cite this work as:
Dombrosky, Jonathan, and Eric Gilmore. 2023. “Year One of
Archaeofaunal Analysis at the Haynie Site (5MT1905).” In
Excavation and Additional Studies at The Haynie Site (5MT1905) by
the Crow Canyon Archaeological Center Annual Report 2022, edited by
Kellam Throgmorton, Susan C. Ryan, Jonathan Dombrosky, Benjamin
Bellorado, Steven Copeland, Kate Hughes, and Jamie Merewether, 45–88. https://www.crowcanyon.org/wp-content/uploads/2023/02/ncop-annual-report-2022-final.pdf.